
ХРЯЩЕВАЯ ТКАНЬ [textus cartilagineus (LNH)] — разновидность соединительной ткани, выполняет опорную функцию.
Xрящевая ткань входит в состав скелета (см.) в виде хрящевых покрытий суставных поверхностей костей (суставной хрящ), хряща межпозвоночных дисков, реберных хрящей, а также формирует внескелетные опорные структуры (хрящи гортани, трахеи, бронхов, хрящевую часть евстахиевой трубы, хрящевые пластинки ушной раковины, носа и др.).
В эмбриогенезе хрящевая ткань образуется из мезенхимы (см.). Предшественниками хондроцитов являются мало-дифференцированные прохондробласты и хондробласты. Они составляют основную массу закладки хрящевой ткани в процессе гистогенеза, а в дальнейшем присутствуют в надхрящнице. На ранних этапах внутриутробного развития почти весь скелет высших позвоночных и человека является хрящевым. В этот период хрящевая ткань составляет до 45% веса (массы) тела. В процессе антенатального и раннего постнатального развития хрящевая ткань в основном замещается костной тканью (см. Кость), в результате чего у взрослого человека масса всех хрящевых образований не превышает 2% массы тела.
Все хрящевые образования за исключением суставного хряща покрыты надхрящницей, состоящей из плотной волокнистой соединительной ткани, богатой сосудами. Надхрящница обеспечивает рост и питание хряща. Кроме того, питание суставного хряща осуществляется при активном участии синовиальной жидкости (см.), находящейся в суставной полости.
Хрящевая ткань состоит из хондроцитов (хрящевых клеток) и хрящевого матрикса. Хондроциты представляют собой крупные клетки овальной или округлой формы с небольшими отростками. В зависимости от степени зрелости различают хондроциты нескольких типов. Хондроцит I типа — молодая активная клетка с высокими показателями синтеза ДНК, обладающая способностью к митозу (см.). Зрелые хондроциты II и III типа содержат в цитоплазме хорошо развитую эндоплазматиче-скую сеть и комплекс Гольджи, активно продуцируют и секретируют коллаген, гликопротеиды, протеогликаны. Для них характерен амитотический тип деления (см. Амитоз).
В зрелом хряще присутствуют также пузырчатые сильно вакуолизированные разрушающиеся клетки, окончившие свой жизненный цикл. Хондроциты находятся в полостях (лакунах) матрикса изолированно или группами, образовавшимися в результате деления одной клетки (изогенные группы). Стенки лакуны представляют собой плотный волокнистый коллагеновый каркас (так наз. перицеллюлярную корзину), который защищает клетки от механических воздействий. Внутри лакуны хондроциты окружены тонкофибриллярным богатым водой основным веществом.
В хрящевом матриксе различают волокна и основное вещество. Волокнистыми компонентами хрящевого матрикса являются коллаген II типа, эластин, белки неколлагеновой природы, гликопротеиды, протеогликаны. Макромолекулы коллагена образуют волокнистые структуры при взаимодействии с гликопротеидами и протеогликанами.
Основное вещество состоит из протеогликанов и гликопротеидов и не является аморфным. Выявлена строгая ориентация, упорядоченность в расположении макромолекул и их агрегатов; векторами ориентации являются как направление волокон коллагена, так и расположение хондроцитов.
Хрящевой ткани свойственна четкая упорядоченность во взаимном расположении клеток и матрикса. В ней принято различать территориальные и межтерриториальные участки. Территориальные участки образованы изогенными группами клеток, окруженных основным веществом и ограниченных волокнистым каркасом циркулярно расположенных коллагеновых волокон. Межтерриториальные участки представлены пучками волокон с прослойками основного вещества, ориентированных соответственно вектору силовых линий распределения нагрузки.
В зависимости от преобладания тех или иных волокнистых компонентов и степени маскировки их гомогенным основным веществом принято различать гиалиновый, волокнистый и эластический хрящи. Наиболее часто в организме встречается гиалиновый хрящ. Гиалиновыми являются суставные и реберные хрящи, а также хрящи носа, гортани (щитовидный и перстневидный), эпифизарный хрящ длинных трубчатых костей, хрящи трахеи и бронхов. Нативный гиалиновый хрящ — плотный, упругий, жемчужно-белый (стекловидный), что связано со значительным содержанием в нем гомогенного основного вещества, богатого протеогликанами, при удалении которых выявляется волокнистый коллагеновый каркас.
Для волокнистого хряща характерно наличие выраженных пучков коллагеновых волокон, а также гетерогенность клеток (наряду с хондроцитами в нем присутствуют и фибробласты). Из волокнистого хряща построены межпозвоночные диски, непрерывные соединения (синхондрозы), а также участки сухожилий и связок в месте их прикрепления.
Эластический хрящ обнаруживается в ушной раковине, надгортаннике, рожковидных и черпаловидных хрящах гортани. Он отличается большим содержанием в матриксе эластических волокон и не подвергается обызвествлению.
Своеобразной разновидностью хрящевой ткани является хондроидная ткань стромы сердца, сохраняющаяся в отдельных участках фиброзных колец у взрослых.
Регенерация хрящевой ткани осуществляется за счет малодифференцированных клеток надхрящницы, а также, по-видимому, благодаря способности хондроцитов при определенных условиях к митотическому делению.
Биохимия хрящевой ткан и химический состав хрящевой ткани в связи с бедностью клетками практически полностью определяется составом ее матрикса, или межклеточного вещества (см.). Хрящевая ткань богата водой (более 70%); сухой остаток составляет около 30%, в нем содержится примерно 50% коллагена (см.), причем специфическим для хрящевой ткани является коллаген II типа, молекулы которого состоят из трех одинаковых полипептидных альфа-цепей. Кроме того, в хрящевой ткани содержится несколько своеобразных, так называемых минорных, коллагенов. В нормальном гиалиновом хряще коллаген II типа составляет основную массу коллагена, в эластическом и фиброзном хрящах наряду с коллагеном II типа присутствует также коллаген I типа. В суставных хрящах концентрация коллагена наиболее высока в поверхностном слое.
Другим компонентом хрящевой ткани являются гликозаминогликаны (см. Мукополисахариды), суммарное содержание которых в эмбриональном периоде достигает 25% сухого остатка, затем постепенно снижается и в пожилом возрасте составляет 14%. Сульфатированные гликозаминогликаны — хондроитинсульфаты (см. Хондроитинсерные кислоты) и кератансульфат — присоединены к так наз. стержневому белку и образуют макромолекулы протеогликанов (протеинполисахаридов, хондромукопротеинов) массой 1 000 000 — 3 000 000. Особенностью хрящевой ткани является то, что протеогликаны соединяются с помощью гиалуроновой кислоты (см. Гиалуроновые кислоты) в агрегаты массой до 50 000 000 — 100 000 000. Агрегаты протеогликанов удерживают в связанном состоянии основную часть содержащихся в хрящевой ткани воды и растворов электролитов, благодаря осмотическому эффекту способствуют поддержанию коллагенового каркаса в расправленном состоянии и обеспечивают диффузию веществ в хрящевую ткань, не содержащую кровеносных сосудов.
Белки неколлагеновой природы составляют 10—20% сухого остатка, в том числе белки, связанные с гликозаминогликанами 7 — 13%, структурные гликопротеиды (см.) и липопротеиды (см.) 3 — 7%, липиды (см.) и дезоксирибонуклеиновые кислоты (см.) 1,3—1,8%. Клетки и матрикс хрящевой ткани содержат также гликоген (см.), а хрящевая ткань эпифизарных отделов костей — фосфорно-кальциевые соли.
Обмен веществ в хрящевой ткани в связи с отсутствием васкуляризации осуществляется хондроцитами (благодаря физическим свойствам и специфическому строению матрикса) и обеспечивается энергией за счет гликолиза (см.), протекающего преимущественно по анаэробному типу. Обмен веществ весьма интенсивен в период роста, особенно в эпифизарных хрящах, но затем резко замедляется, и зрелая хрящевая ткань характеризуется выраженной метаболической инертностью. Xрящевая ткань обладает способностью к обратимой деформации в условиях значительных механических нагрузок, а также слабой иммунологической реактивностью в связи со способностью гидратированного матрикса задерживать и изолировать антигены.
В процессе старения организма в хрящевой ткани уменьшается концентрация протеогликанов, а следовательно, и стелень гидратированности матрикса.
В хондроцитах накапливаются гликоген и липиды, уменьшаются размеры комплекса Гольджи (см. Гольджи комплекс) и эндоплазматической сети (см. Эндоплазматический ретикулум), а также число митохондрий (см.). Клетки вакуолизируются и гибнут, а лакуны заполняются основным веществом. В матриксе откладываются соли кальция и уменьшается содержание воды, что ведет к потере хрящом эластодинамических свойств.
Читайте также: Преимущества одежды из трикотажа по сравнению с одеждой из тканей
В патологических условиях обмен веществ в хрящевой ткани нарушается: повышается активность протеолитических ферментов, интенсифицируются катаболические и биосинтетические процессы, происходит нарушение структуры и агрегации протеогликанов, появляются несвойственные хрящевой ткани коллагены, отмечаются отложение пигментов и избыток липидов.
Библиогр.: Жаденов И. И. и Пастель В. Б. Обменные процессы в суставном хряще в норме (возрастной аспект) и при патологии (остеоартроз), Ортоп. и травмат., № 3, с. 65, 1982, библиогр.; Многотомное руководство по ортопедии и травматологии, под ред. Н. П. Новаченко, т. 1, с. 427, 606, М., 1967; Многотомное руководство по патологической анатомии, под ред. А. И. Струкова, т. 5, с. 234, 433, М., 1959; Павлова В. Н. Синовиальная среда суставов, с. 155, М., 1980; Рейнберг С. А. Рентгенодиагностика заболеваний костей и суставов, кн. 1, М., 1964; Слуцкий Л. И. Биохимия нормальной и патологически измененной соединительной ткани, Л., 1969; Тагер И. Л. Рентгенодиагностика заболеваний позвоночника, с. 101, М., 1983; Франке К. Спортивная травматология, пер. с нем., с. 74, М., 1981; Хэм А. и Кормак Д. Гистология, пер. с англ., т. 3, М., 1983; KneseK. -H. Stiitzgewebe und Skelett-system, В. u. а., 1979; Thompson R. С. a. Robinson H. J. Articular cartilage matrix metabolism, J. Bone Jt. Surg. v. 63-A, p. 327, 1981.
В. H. Павлова (ан., гист., эмбр.), Л. И. Слуцкий (биохим.).
Матрикс хрящевой ткани это
Хрящ — это соединительная ткань, состоящая из клеток (хондробластов и хондроцитов), которые продуцируют внеклеточный матрикс, включающий в себя протеогликаны и волокна коллагена с большим содержанием воды. Прочность хрящей при растяжении обеспечивается именно коллагеном. Своей устойчивости к давлению хрящ обязан способности протеогликана притягивать и удерживать воду.
Выделяют несколько типов хрящей: суставной или гиалиновый; фиброзный или волокнистый, встречающийся в местах прикрепления к костям связок и сухожилий; эластичный, из него состоят мениски и межпозвоночные диски; и хрящ зоны роста, расположенный в местах формирования костной ткани (окостенения).
С возрастом содержание воды в хряще и количество связей между молекулами коллагена постепенно снижаются. В результате хрящевая ткань становится менее эластичной и хуже сопротивляется растяжению, скручиванию и компрессионным нагрузкам. Другими словами, с возрастом хрящ становится более уязвимым для повреждений.
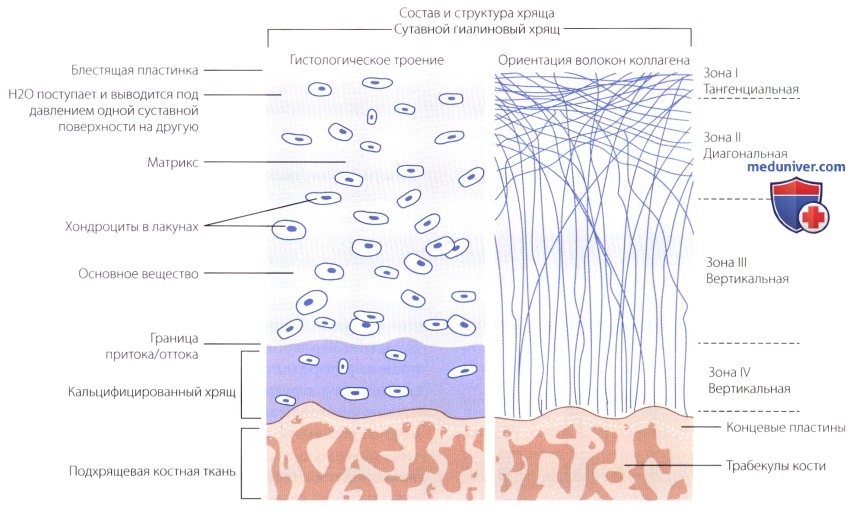
Суставной хрящ образует синовиальную поверхность суставов. Он соединяется с нижележащей костью посредством встречно-гребенчатой конструкции, напоминающей детскую мозаику. Регенерация этого хряща происходит очень медленно и несопоставима со сроками восстановления других тканей сустава. После повреждения гиалиновый хрящ может замещаться волокнистым хрящом, механические свойствами которого хуже.
В суставном хряще отсутствуют кровеносные сосуды, поэтому его питание зависит исключительно от нагрузки сустава. При этом через пористый верхний слой в хрящевой матрикс поступают растворимые в воде питательные вещества и удаляются продукты обмена.
Эластичный хрящ межпозвоночных дисков допускает лишь минимальное движение между соседними позвонками, обеспечивая амортизацию. В связи с определенной ориентацией волокон, диски подвержены повреждениям при сгибании и скручивании. Эластичный хрящ также представлен в менисках коленного сустава. Здесь его функцией является не только амортизация при ударах, но и увеличение рабочей поверхности сустава, что обеспечивает дополнительную стабильность.
Благодаря эластину, входящему в состав эластичного хряща, последний способен восстанавливать свою первоначальную форму после деформации.
Видео гистология хряща (хрящевой ткани)
Редактор: Искандер Милевски. Дата публикации: 27.11.2021
Что такое внеклеточный матрикс и почему его все изучают
Внеклеточный матрикс — неотъемлемая часть любой ткани.
Автор
Редакторы
Внеклеточный матрикс (ВКМ) — многокомпонентная субстанция, в которую погружены все клетки нашего организма. В последнее десятилетие интерес к внеклеточному матриксу значительно возрос. Это связано с установлением его роли в старении, клеточной дифференцировке, успешной терапии рака и лечении некоторых наследственных заболеваний. Мы подготовили цикл статей, в котором расскажем об организации внеклеточного матрикса, болезнях, связанных с его патологиями, роли ВКМ в старении и подходах к корректировке возрастных изменений.
В первой статье цикла мы рассказываем о компонентах и функциях внеклеточного матрикса, разбираемся, какую практическую пользу может принести его изучение, а также вкратце освещаем самые важные открытия в этой области, совершенные за последний год.
Редакция благодарит Татьяну Голубеву за экспертные консультации и комментарии при подготовке статьи.
Что такое внеклеточный матрикс?
Внеклеточный матрикс (ВКМ, extracellular matrix, ECM) объединяет разрозненные клетки в единый многоклеточный организм. ВКМ похож на государство, в котором живут граждане-клетки: он отвечает за благополучие клеток и управляет всеми аспектами их жизни — от питания и размножения до смерти. Для клеток ВКМ — это и квартира, и магазин, и интернет, и общественный транспорт.
ВКМ: основные игроки
Создатели ВКМ. Компоненты ВКМ синтезируют специализированные клетки. В соединительной ткани наиболее распространены фибробласты, однако создавать ВКМ умеют и другие клетки: в хрящах, например, это хондроциты, а в костях — остеобласты. Компоненты матрикса могут синтезировать и клетки прилегающих органов: например, клетки эпителия сосудов производят компоненты рыхлой соединительной ткани.
Разрушители ВКМ. Порядок в ВКМ наводят белки — разрушители его компонентов. Наиболее важные из них, металлопротеиназы, «расчищают путь» клеткам, которые движутся в ВКМ, и уничтожают старые и «сломанные» компоненты матрикса.
Основное вещество ВКМ. Основу матрикса формируют гиалуроновая кислота и особые белки: гликопротеины и протеогликаны (рис. 1). В состав ВКМ входит и множество других белков со специфической функциональной нагрузкой — интегрины, фибронектины, эластин, — о них мы поговорим в следующем разделе.
Тем, кто хочет узнать больше о построении и разрушении ВКМ, советуем почитать замечательный обзор на эту тему — «Кто рубит коллагеновый лес» [1].
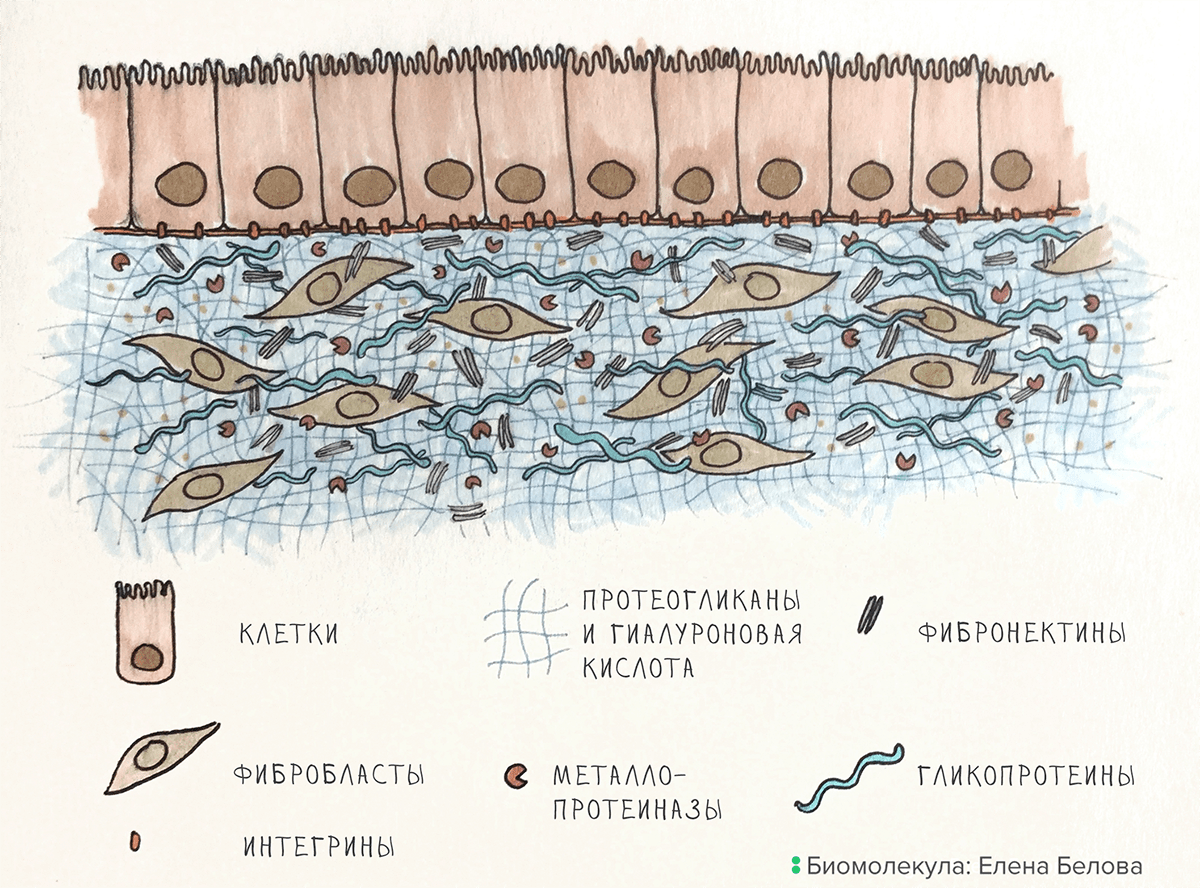
Рисунок 1. Организация ВКМ на примере кожи. Фибробласты создают ВКМ, металлопротеиназы его разрушают. Клетки эпидермиса связаны с ВКМ при помощи интегринов.
Компоненты ВКМ
Выделяют два подкласса белков, содержащих углеводы, — протеогликаны и гликопротеины. Оба подкласса входят в состав ВКМ, однако между ними есть существенные различия.
В гликопротеинах доля углеводов не превышает 20%, углеводные цепи короткие, имеют нерегулярное строение и не содержат уроновых кислот.
К гликопротеинам относятся такие важные структурные белки, как коллаген и эластин. За счет самого распространенного структурного белка в организме — коллагена — ВКМ приобретает прочность, а за счет эластина — гибкость и эластичность.
Протеогликаны — сложные белки с высокой степенью гликозилирования, часто имеющие в своем составе уроновые кислоты. В отличие от гликопротеинов, около 90–95% массы такой молекулы составляют длинные углеводные цепи регулярного строения, а на белки приходится лишь 5–10%. Такое строение обуславливает высокую молекулярную массу протеогликанов. Например, у аггрекана, белка хрящевой ткани, она может достигать 5 МДа [2]! Протеогликаны запасают воду и полезные вещества, то есть работают как «магазин», из которого клетки получают необходимые «продукты». Например, они могут фиксировать ионы кальция в очагах оссификации. Кроме того, они играют роль межтканевых прослоек и смазочного материала в суставах.
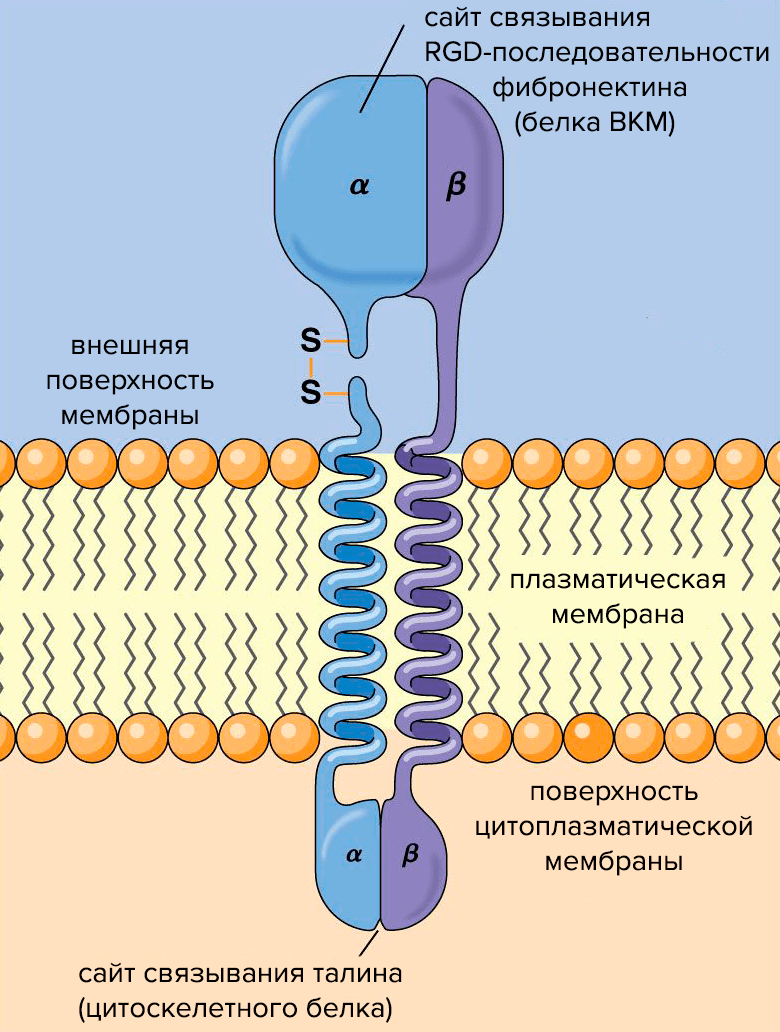
Рисунок 2. Схема строения молекулы интегрина. Субъединицы интегрина (α и β) пронизывают плазматическую мембрану, связывая клетку с внешней средой.
Читайте также: Легкая плотная хлопчатобумажная ткань с рельефным тканым рисунком 7 букв
Интегрины — трансмембранные клеточные рецепторы, которые взаимодействуют с ВКМ и участвуют в межклеточной коммуникации. В результате этих взаимодействий в клетках активируются сигнальные каскады, регулирующие экспрессию генов, отвечающие за пролиферацию и дифференцировку клеток, их выживание или апоптоз. То есть система интегрины—ВКМ — нечто среднее между «интернетом» и «рабочим чатом».
Структурно интегрины представляют собой облигатные гетеродимеры: каждый состоит из одной α- и одной β-субъединицы (рис. 2). Альфа-субъединицы определяют специфичность интегрина к лиганду — молекуле, способной образовывать комплекс с интегрином и таким образом воздействовать на клетку. Бета-субъединицы связаны с компонентами цитоскелета и обеспечивают передачу сигнала в клетке. О работе интегринов подробно рассказано в статье «Клетки под давлением» [3].
Интегрины улавливают химические и физические сигналы из внеклеточного матрикса и проводят их в клетку. Сигнал от интегринов передается в ядро через белки цитоскелета и сигнальные белки — так ВКМ управляет экспрессией генов и регулирует клеточную пролиферацию. При посредничестве белков цитоскелета ВКМ также управляет формой и движениями клеток.
Фибронектины — это белки клеточной адгезии. Растворимые предшественники фибронектинов синтезируюся внутри клеток и затем попадают в ВКМ. Интегрины превращают фибронектины в нерастворимые нити — компоненты внеклеточного матрикса [4]. Они помогают клеткам закрепляться на поверхностях и отвечают за их рост и перемещение в ВКМ [5].
Гиалуроновая кислота (ГК) синтезируется встроенными в мембрану белками и затем «выдавливается» через нее в межклеточное пространство. По составу ГК похожа на углеводную часть протеогликана и представляет собой полимер из остатков D-глюкуроновой кислоты и D-N-ацетилглюкозамина. ГК помогает интегринам проводить сигналы в клетку, регулирует клеточный ответ на эти сигналы и, подобно фибронектину, дает клеткам возможность закрепляться на различных поверхностях [6]. Образно говоря, ГК выполняет задачи «интернет-провайдера» и «билета на общественный транспорт».
Почему так важно изучать внеклеточный матрикс?
Внеклеточный матрикс присутствует во всех тканях организма, поэтому сбои в его функционировании ведут к развитию болезней соединительной ткани, преждевременному старению и гибели клеток. Самый очевидный стимул для изучения ВКМ — необходимость лечения заболеваний, связанных с нарушениями структуры соединительной ткани. Таких болезней много, они могут протекать тяжело и значительно ухудшать качество жизни пациентов. Вот несколько примеров.
Мутации в генах, отвечающих за синтез структурных белков ВКМ, приводят к врожденным патологиям [7]. Поскольку соединительная ткань — основа всех систем организма, пострадать от генетических нарушений может любой орган. Такие нарушения приводят:
- на уровне костей — к несовершенному остеогенезу,
- на уровне кожи — к синдрому Элерса—Данлоса, при котором кожа становится слишком эластичной,
- на уровне мышечной ткани — к врожденной мышечной дистрофии (CMD).
Из всех типов генетических нарушений ВКМ лучше всего изучена врожденная миодистрофия. Это заболевание развивается из-за нескольких мутаций, нарушающих работу мышц. В результате мутации в гене ITGA7, кодирующем интегрины мышечных клеток, нарушается связь клеток с ВКМ. Это гибельно для мышечной ткани: потеря связей с ВКМ запускает апоптоз — программируемую смерть клеток.
Преждевременное разрушение ВКМ — еще одна серьезная проблема. Здоровый внеклеточный матрикс постоянно обновляется и реструктурируется. За это отвечает семейство металлопротеиназ, белков — разрушителей ВКМ.
Металлопротеиназы — ферменты, в каталитический центр которых входят ионы металлов, в основном цинка. Отсюда и название с приставкой «металло-». Помимо каталитического центра, который отвечает за разрушение ВКМ, у металлопротеиназ есть регуляторный продомен, оберегающий фермент от преждевременной активации и ложного срабатывания.
Как именно металлопротеиназы разрушают коллаген и «отцепляют» клетки от волокон внеклеточного матрикса, рассказано в статье «Кто рубит коллагеновый лес» [1]. Сейчас же нам важно понять, что если металлопротеиназы «выходят из-под контроля», то они начинают буквально сметать всё на своем пути. Бесконтрольное разрушение ВКМ приводит к фиброзу и может вызвать рак [8] — например, рак предстательной железы [9].
Еще одна патология, вызываемая нарушениями в процессах разрушения и формирования ВКМ, — болезнь Крона (хроническое воспаление кишечника) [10]. По мере ее развития возникает фистулизация и фибростеноз кишечника (рис. 3).
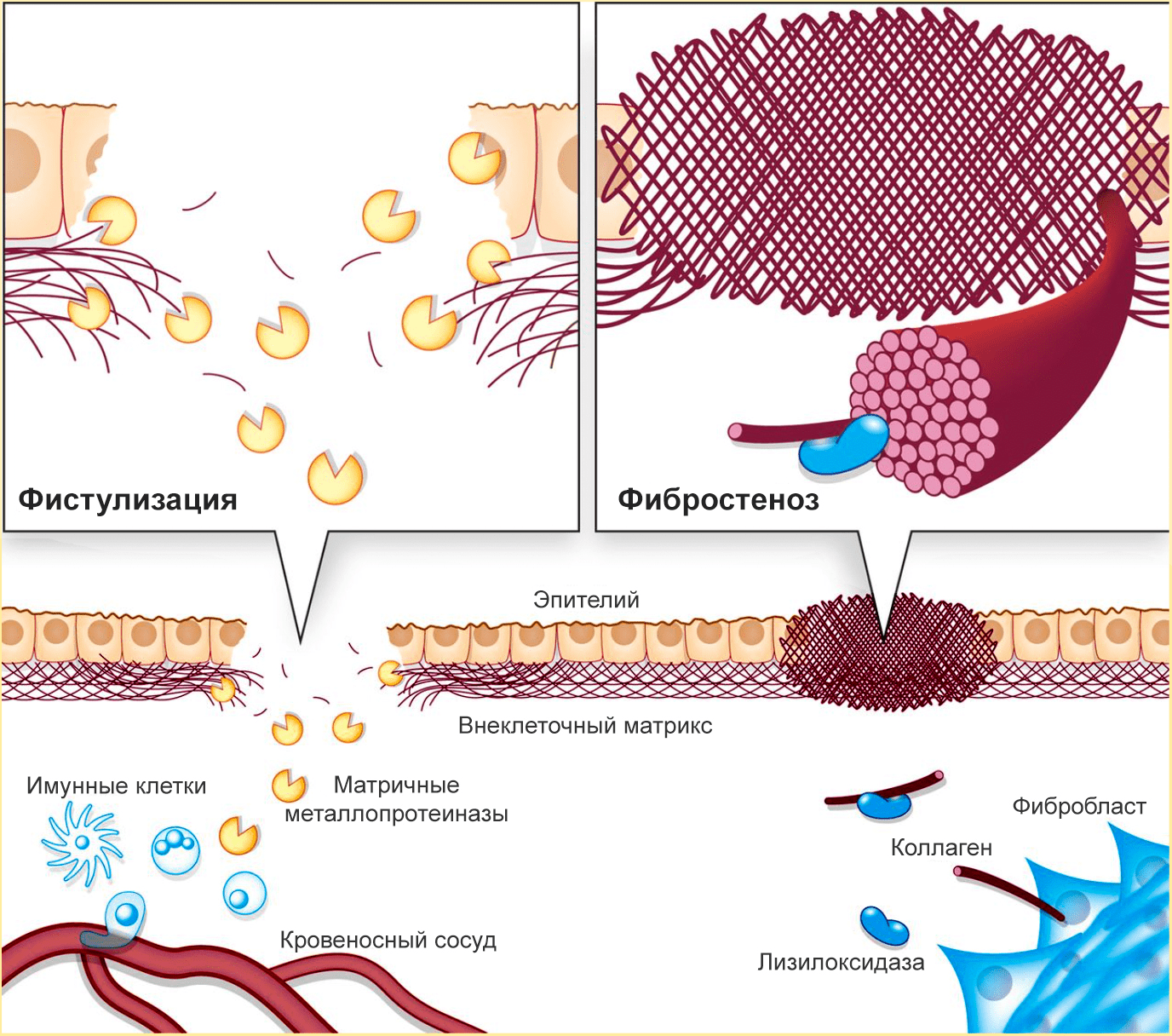
Рисунок 3. При болезни Крона происходит фистулизация и фибростеноз кишечника. Из-за избыточного разрушения ВКМ в кишечной стенке образуются отверстия, а из-за избыточного синтеза коллагена сужается просвет кишечника.
Фистулизация — образование отверстий в кишечной стенке. Этот процесс связан с неконтролируемой активностью металлопротеиназ, разрушающих коллаген слизистой оболочки кишечника. Так как целостность слизистой нарушается, иммунные клетки из кровеносных сосудов проникают в кишечную стенку — так развивается воспаление, а через некоторое время появляется отверстие в стенке кишечника.
Фибростеноз — сужение кишечника. Реагируя на повреждения, фибробласты интенсивно производят коллаген. Затем активируется лизилоксидаза — внеклеточный медьсодержащий фермент, катализирующий образование сложных поперечный связей в коллагене и эластине. Лизиолоксидаза необходима для формирования зрелых коллагеновых волокон, но при ее избытке начинаются проблемы. Этот фермент создает прочную коллагеновую сеть, «запечатывающую» поврежденные кишечные стенки, но из-за избыточной жесткости коллагеновых «печатей» фиброз только усиливается. В результате возникают воспаление и (иногда) непроходимость кишечника. Фистулизация усиливает фибростеноз: неуправляемое разрушение коллагена стимулирует его столь же неуправляемый синтез.
Избыточный синтез ВКМ часто говорит о том, что у человека рак и у этого рака плохой прогноз [11]. Кроме того, сам ВКМ может способствовать росту опухоли и распространению метастазов — это доказано для опухоли головного мозга. Особенно неприятно, что существующие методы лечения — например, лучевая терапия — способны заставить ВКМ послать клеткам сигнал, который может привести к рецидиву опухоли мозга [12].
Состав ВКМ головного мозга уникален: в нём очень много гиалуроновой кислоты и при этом гораздо меньше коллагена, фибронектина и других компонентов, характерных для ВКМ всех остальных тканей. Несмотря на важность ГК для нормальной работы тканей, в некоторых случаях она помогает проводить в клетки опухоли сигналы, делающие их более агрессивными. О том, как именно ГК «дразнит» раковые клетки, мы поговорим в разделе «Что нового мы узнали о внеклеточном матриксе за последний год?»
Лучевая терапия стимулирует работу мембранных белков HAS2, которые отвечают за синтез ГК. В результате они синтезируют больше гиалуроновой кислоты, а чем больше ГК, тем агрессивнее становятся опухолевые клетки. Получается, что лучевая терапия может стимулировать метастазирование опухоли. Таким образом, из-за особенностей ВКМ лучевая терапия помогает только на время: опухоль часто возвращается, становясь еще более опасной. Отчасти по этой причине некоторые виды опухолей мозга так плохо поддаются лечению.
Проблемы с ВКМ — важная причина старения
Первые признаки старения, которые сразу бросаются в глаза, — слабость, хрупкость костей, появление морщин и старческих пятен. Многие из этих проблем связаны с необратимыми изменениями во внеклеточном матриксе.
Одна из причин старения кожи — нарушение работы фибробластов (клеток, синтезирующих компоненты внеклеточного матрикса). При этом благополучие фибробластов зависит от состояния внеклеточного матрикса — получается замкнутый круг.

Рисунок 4. Микрофотография коллагеновых нитей в коже. а — «Целый» коллаген в молодой коже. б — Фрагментированный коллаген в стареющей коже. Стрелки указывают на старые, «разорванные» на кусочки нити коллагена.
Читайте также: Камбий конус нарастания стебля какая ткань
Чтобы фибробласты хорошо себя чувствовали, им нужно цепляться за интактные, целые коллагеновые нити. Но со временем эти нити фрагментируются, и фибробластам становится не к чему крепиться, чтобы создать новый, целый коллаген (рис. 4) [13]. Если бы мы научились разрывать этот замкнутый круг, то нам, вполне возможно, больше никогда не понадобились бы антивозрастные услуги косметологов и пластических хирургов.
Старение костной ткани чаще всего связано с нарушением работы остеобластов. Эти клетки создают костный ВКМ, синтезируя коллаген и особые гликопротеины, которые участвуют в первом этапе минерализации коллагена. На втором этапе коллаген превращается в прочный внеклеточный матрикс — основу кости [14].
С течением лет старые остеобласты погибают, а новые делятся хуже и хуже. Оставшимся в живых стареющим клеткам становится всё сложнее справляться с созданием внеклеточного матрикса. Из-за этого кости у пожилых людей становятся очень хрупкими и плохо заживают после переломов.
Если бы мы могли заставить остеобласты делиться или хотя бы выяснили, как помочь им эффективнее минерализовать соединительную ткань, у пожилых людей появилась бы возмножность быстрее восстанавливаться после переломов. Такие работы уже ведутся!
В одной из подобных работ исследователи взяли коллагеновый гель, добавили к нему два неколлагеновых белка, ответственных за минерализацию коллагена (остеокальцин и остеопонтин), и создали на их основе искусственный внеклеточный матрикс. Этот исусственный ВКМ исследователи «предложили» остеобластам, полученным из стволовых клеток костного мозга. В результате активизировалось деление остеобластов, и эти остеобласты начали производить больше компонентов внеклеточного матрикса — правда, пока только в лабораторных условиях [14]. Чтобы искусственный ВКМ появился в кабинете травматолога и оказал содействие в восстановлении костей реальных пациентов, потребуется провести еще много исследований.
Что нового мы узнали о внеклеточном матриксе за последний год?
Над изучением ВКМ работают сотни исследовательских групп по всему миру, и каждый день появляется множество публикаций по этой теме. Давайте рассмотрим несколько свежих работ, чтобы получить представление о ключевых направлениях современных исследований. Вот что мы узнали о компонентах ВКМ в 2018 году.
Металлопротеиназы
Когда металлопротеиназы преждевременно разрушают эластин и коллаген легких, возникает соединительнотканное заболевание легких — эмфизема. Ученые давно стремились узнать, какой генетический компонент влияет на чрезмерную активность металлопротеиназ при этом заболевании.
Исследователи из немецкого Общества имени Макса Планка [15] связали чрезмерную активность металлопротеиназ в легких со снижением активности гена Myh10. Если этот ген «отключали» в мышиных легких, их внеклеточный матрикс формировался неправильно, разрушались межальвеолярные перегородки, с укрупнением альвеол сокращалась суммарная площадь их поверхности, а значит, должен был страдать газообмен. То есть события развивались по сценарию, типичному для человеческой эмфиземы лёгких. Кроме того, авторы работы выявили снижение экспрессии гена MYH10 в легких людей, страдающих эмфиземой.
Не исключено, что в будущем мы научимся управлять процессами, которые протекают в матриксе, используя редактирование генома и генную терапию.
Гликопротеины и металлопротеиназы
Яркий пример коварства металлопротеиназ — образование аневризм. Так, при аневризме брюшной аорты матриксные металлопротеиназы разрушают компоненты, из которых строится ВКМ этого крупного сосуда. Вскоре эта область воспаляется благодаря деятельности иммунных клеток — макрофагов. Механизмы, заставляющие металлопротеиназу разрушать аорту, долгое время оставались неизвестными.
Американским исследователям удалось выяснить, что макрофаги синтезируют нетрин-1 — белок, активирующий клетки гладких мышц сосудов [16]. Под воздействием нетрина-1 гладкомышечные клетки активируют свободные металлопротеиназы, разрушающие соединительную ткань сосуда.
Возможно, когда-нибудь мы научимся использовать иммунные клетки, чтобы управлять разрушением внеклеточного матрикса.
Интегрины и фибронектины
Южноафриканские исследователи обнаружили, что на синтез и деградацию фибронектина влияет внутриклеточный белок теплового шока Hsp90. Этот белок воздействует на фибронектин посредством рецептора LRP1 [4]. Если заблокировать этот рецептор, количество фибронектина, который накапливается во внеклеточном матриксе, уменьшается. И это очень хорошо — ведь из-за избыточного накопления фибронектина развиваются многие патологии ВКМ.
Не исключено, что если мы обнаружим подобные рецепторы в клетках разных тканей и научимся на них воздействовать, то сможем предотвращать болезни, связанные с накоплением ВКМ — например, фиброз легких.
Гиалуроновая кислота
Как компонент внеклеточного матрикса гиалуроновая кислота участвует в передаче сигнала от ВКМ в клетку и даже может превратить здоровую клетку в злокачественную [17]. Гиалуроновая кислота воздействует на клетки через рецептор CD44.
Удалось выяснить, что интенсивность сигнала, запускающего злокачественный процесс в клетке и определяющего, насколько опасна будет получившаяся раковая клетка, зависит от концентрации гиалуроновой кислоты и от ее молекулярной массы.
Гиалуроновая кислота управляет раковыми клетками, связываясь с трансмембранным белком — интегрином CD44. Активация рецептора CD44 подавляет апоптоз, из-за чего клетка становится «бессмертной», то есть раковой. Чем больше гиалуроновой кислоты, тем больше активируется рецепторов CD44 и тем, соответственно, опаснее и агрессивнее будут раковые клетки.
Если мы научимся управлять гиалуронидазами — ферментами, которые отвечают за модификацию и деградацию гиалуроновой кислоты, — мы сможем предотвращать развитие рака и появление метастазов.
Как создавать ВКМ
Когда мы усовершенствуем технологии создания искусственного ВКМ, мы сможем производить полноценные ткани и использовать их в регенеративной медицине. Работы в этом направлении ведутся уже давно, и в прошлом году удалось сделать несколько важных шагов на пути к технологии искусственного ВКМ.
Исследователи уже научились создавать рабочие 3D-модели ВКМ (рис. 5). В отличие от традиционных 2D-моделей, которые, по сути, представляли собой клеточную культуру в чашке Петри, 3D-модели позволяют создать работающий объемный «макет» нужной нам ткани [18].
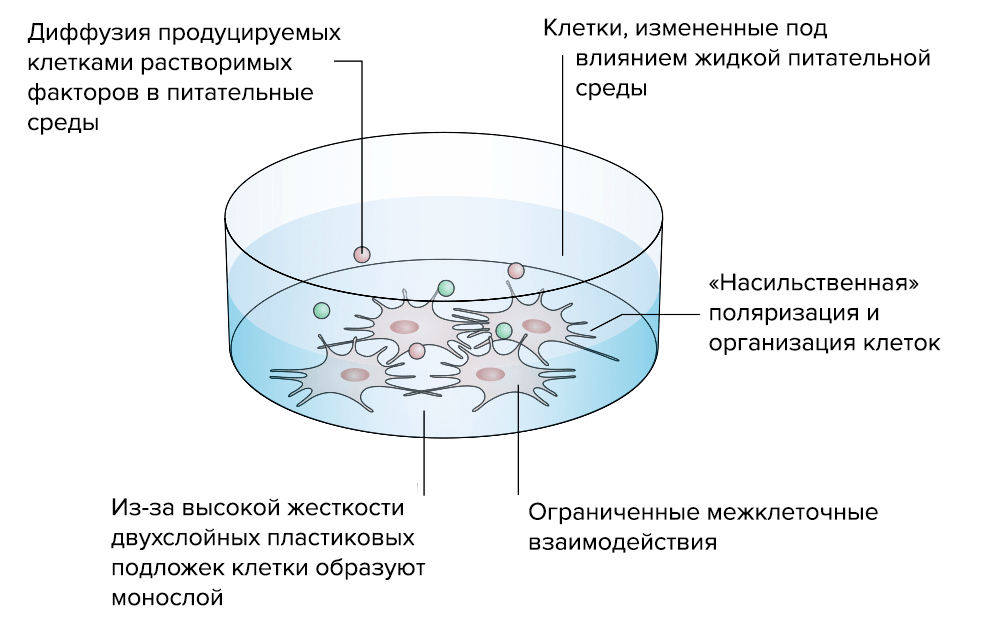
Рисунок 5а. Преимущества «объемной» 3D-модели соединительной ткани перед стандартной «плоской» 2D-моделью. 2D-модель. Клетки, выращенные на двумерной пластиковой подложке, ведут себя неестественно: нарастают в один слой, из-за чего нарушается межклеточное взаимодействие.
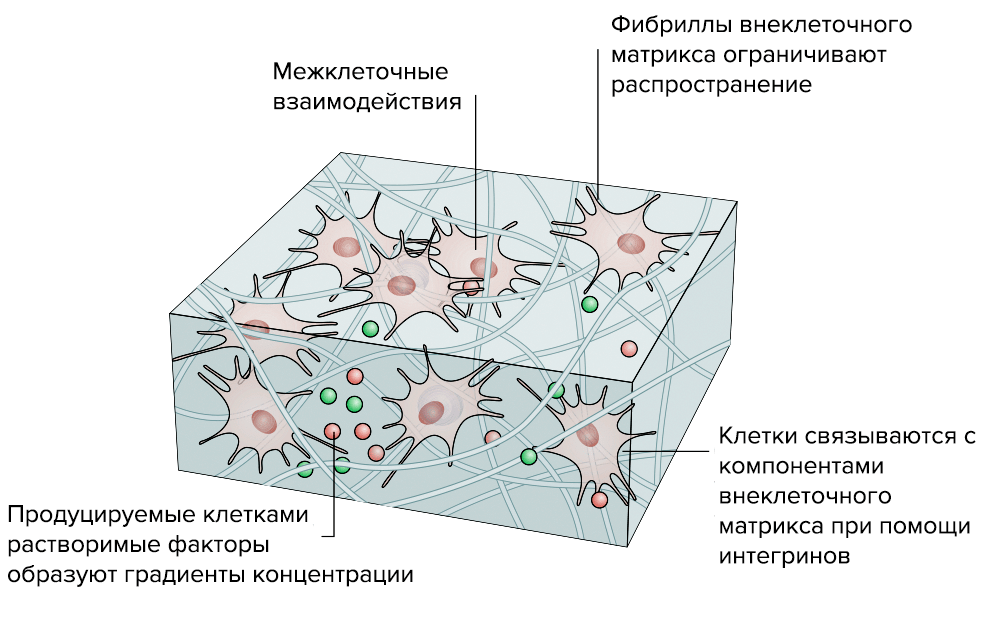
Рисунок 5б. Преимущества «объемной» 3D-модели соединительной ткани перед стандартной «плоской» 2D-моделью. 3D-модель. «Живой макет» ведет себя почти так же, как функциональная ткань: клетки растут и располагаются так, как им «нравится», поэтому у них сохраняется способность к полноценному межклеточному взаимодействию.
3D-модели можно модифицировать и дорабатывать. Уже созданы первые гидрогели — биосовместимые синтетические полимеры, способные удерживать воду. С появлением гидрогелей появилась и теоретическая возможность печатать внутренние органы.
Перед тем как первые напечатанные в лаборатории 3D-органы поступят в больницу, ученым придется решить еще множество проблем. Например, исследователям только предстоит разобраться с тем, как «подвести» к искусственным органам кровеносные сосуды. В этом тоже может помочь понимание биологии ВКМ — ведь мы уже знаем, что внеклеточный матрикс управляет в том числе и ростом сосудов [19].
Заключение
Внеклеточный матрикс — огромная тема, которую очень сложно охватить в одной статье. Ясно одно: если мы сумеем разобраться, как ВКМ функционирует и влияет на клетки, медицина сделает огромный шаг вперед.
Новые открытия в области изучения ВКМ помогут нам:
- победить опасные наследственные болезни,
- эффективно бороться с некоторыми видами рака,
- создавать искусственные органы и ткани,
- возвращать людям молодость и красоту.
Судя по всему, ждать прорыва в изучении ВКМ осталось недолго.