
Механизмы переноса углекислого газа от тканей кровью
С транспортом двуокиси углерода кровью возникает намного меньше проблем, чем с транспортом кислорода, поскольку даже в самых нетипичных условиях двуокись углерода может транспортироваться в гораздо больших количествах, чем кислород. Но количество двуокиси углерода в крови во многом связано с кислотно-щелочным равновесием в жидкостях тела. В нормальных условиях в покое из тканей в легкие транспортируется в среднем 4 мл двуокиси углерода в 100 мл крови.
а) Химические формы транспорта двуокиси углерода. В начале процесса транспорта двуокись углерода диффундирует из клеток ткани в растворенном виде. При входе в тканевые капилляры двуокись углерода включается в ряд быстропротекающих физических и химических реакций (для облегчения понимания просим вас изучить рисунок ниже), необходимых для ее транспорта.
![]()
Транспорт двуокиси углерода кровью
б) Транспорт двуокиси углерода в растворенном виде. Небольшая часть двуокиси углерода транспортируется в легкие в растворенном виде. Вспомните, что PCO2 в венозной крови составляет 45 мм рт. ст., а в артериальной крови — 40 мм рт. ст. При PCO2, равном 45 мм рт. ст., объем двуокиси углерода, растворенный в жидкой части крови, равен примерно 2,7 мл/дл (2,7 об%), а при PCO2, равном 40 мм рт. ст., — 2,4 мл/дл. Разница в объеме растворенной двуокиси углерода между артериальной и венозной кровью составляет 0,3 мл/дл. Таким образом, для выделения в легких транспортируется в растворенном виде только 0,3 мл двуокиси углерода в 100 мл крови. Это составляет около 7% всего объема двуокиси углерода, транспортируемого кровью в нормальных условиях.
в) Транспорт двуокиси углерода в виде иона бикарбоната. Реакция двуокиси углерода с водой в эритроцитах. Влияние карбоангидразы. Растворенная в крови двуокись углерода реагирует с водой, образуя угольную кислоту. Из-за медленного протекания эта реакция не имела бы особого значения, если бы в этом не принимала бы участие находящаяся в эритроцитах карбоангидраза — фермент, который катализирует реакцию между двуокисью углерода и водой, ускоряя ее примерно в 5000 раз, поэтому данная реакция, которая в плазме крови происходит за несколько секунд или минут, в эритроцитах протекает с такой скоростью, что почти полное равновесие достигается за долю секунды. Это позволяет внушительному количеству двуокиси углерода реагировать с водой в эритроците еще до того, как кровь покидает тканевые капилляры.
г) Диссоциация угольной кислоты в ионы бикарбоната и водорода. За еще одну долю секунды образующаяся в эритроцитах угольная кислота (H2CO3) диссоциирует на ионы водорода и бикарбоната (Н+ и HCO3 — ). После этого большинство ионов Н+ присоединяются в эритроцитах к гемоглобину, который является мощным кислотно-щелочным буфером. В свою очередь, многие ионы бикарбоната диффундируют из эритроцитов в плазму, откуда в эритроцит возвращаются ионы хлора. Это обеспечивается наличием специального белка — переносчика ионов бикарбоната и хлора в мембране эритроцитов, который с большой скоростью транспортирует эти ионы в противоположные стороны. Содержание ионов Сl- в эритроцитах венозной крови оказывается больше, чем в эритроцитах артериальной крови. Это явление называют хлорным сдвигом.
Обратимая комбинация двуокиси углерода с водой в эритроцитах с участием карбоангидразы обеспечивает около 70% транспорта двуокиси углерода из тканей в легкие. Таким образом, этот путь транспорта двуокиси углерода является наиболее важным. Действительно, если подопытному животному ввести ингибитор карбоангидразы (ацетазоламид) и таким образом блокировать действие карбоангидразы в эритроцитах, то выведение двуокиси углерода из тканей снижается настолько, что PCO2 в тканях может подниматься до 80 мм рт. ст. вместо нормальных 45 мм рт. ст.
д) Транспорт двуокиси углерода в связи с гемоглобином и протеинами плазмы. Карбогемоглобин. Кроме реакции с водой, двуокись углерода напрямую реагирует с аминными радикалами молекулы гемоглобина, образуя карбаминогемоглобин (CO2Нb). Эта реакция обратима, образующиеся связи слабы, и двуокись углерода легко высвобождается в альвеолах, где PCO2 ниже, чем в капиллярах легких.
Небольшое количество двуокиси углерода образует в капиллярах легких такие же соединения с белками плазмы. Для транспорта двуокиси углерода это не имеет большого значения, т.к. количество таких белков в плазме в 4 раза меньше, чем количество гемоглобина.
Количество двуокиси углерода, которое может переноситься из периферических тканей в легкие при помощи карбаминных связей с гемоглобином и белками плазмы, составляет примерно 30% общего количества двуокиси углерода, транспортируемого кровью, — в норме около 1,5 мл двуокиси углерода в 100 мл крови. Однако учитывая, что эта реакция протекает намного медленнее, чем реакция двуокиси углерода с водой в эритроцитах, сомнительно, чтобы в нормальных условиях при помощи карбаминного механизма переносилось более чем 20% общего количества транспортируемой двуокиси углерода.
Читайте также: Модель в составе ткани что это
Видео физиология газообмена в легких и транспорта газов кровью — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Перенос углекислого газа кровью от тканей к легким
В организме человека, не выполняющего физической работы (состояние покоя), от тканей к легким каждую минуту переносится примерно 180 мл углекислого газа. Эту величину легко рассчитать. Если дыхательный коэффициент равен 0,85, то при поглощении тканями в покое 200 мл кислорода в минуту должно образовываться около 170 мл углекислого газа (200•0,85). На самом деле величина несколько больше, поскольку количество поглощаемого в покое кислорода колеблется от 200 до 240 мл в минуту.
В целом за сутки с вдыхаемым воздухом в организм человека поступает примерно 600 л кислорода и выделяется в окружающую среду 480 л углекислого газа (примерно 942,8 г), что соответствует 21,4 моль углекислого газа.
Организм располагает несколькими механизмами переноса СО2 от тканей к легким. Часть его переносится в физически растворенном виде. Растворимость СО2 в плазме крови в 40 раз превышает растворимость в ней кислорода, тем не менее при небольшой артериовенозной разнице РСО2 (напряжение СО2 в венозной крови, притекающей к легким по легочной артерии, равно 60 гПа, а в артериальной крови – 53,3 гПа) в физически растворенном виде может быть перенесено в покое 12–15 мл СО2, что составляет 6–7% от всего количества переносимого углекислого газа.
Некоторое количество СО2 может переноситься в виде карбаминовой формы. Оказалось, что СО2 может присоединяться к гемоглобину посредством карбаминовой связи, образуя карбгемоглобин, или карбаминогемо-глобин (впервые мысль о наличии углекислого газа, непосредственно связанного с гемоглобином, была высказана И.М. Сеченовым):
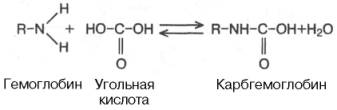
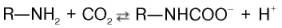
Карбгемоглобин – соединение очень нестойкое и чрезвычайно быстро диссоциирует в легочных капиллярах с отщеплением СО2.
Количество карбаминовой формы невелико: в артериальной крови оно составляет 3 об. %, в венозной – 3,8 об. % . В виде карбаминовой формы из ткани к легким переносится от 3 до 10% всего углекислого газа, поступающего из тканей в кровь. Основная масса СО2 транспортируется с кровью к легким в форме бикарбоната, при этом важнейшую роль играет гемоглобин эритроцитов.
Как отмечалось, кислотный характер оксигемоглобина выражен значительно сильнее, чем гемоглобина (константа диссоциации ННbО2 примерно в 20 раз больше константы диссоциации ННb). Важно также запомнить, что поступающий в ткани с кровью оксигемоглобин является более сильной кислотой, чем Н2СО3, и связан с катионом калия. Эту калийную соль оксигемоглобина можно обозначить как КНbО2 (рис. 17.7). В периферических капиллярах большого круга кровообращения гемоглобин эритроцитов отдает кислород тканям (КНbО2 —> О2 + KHb), его способность связывать ионы водорода увеличивается. Одновременно в эритроцит поступает продукт обмена – углекислый газ. Под влиянием фермента карбоангидразы углекислый газ взаимодействует с водой, при этом образуется угольная кислота. Возникающий за счет угольной кислоты избыток водородных ионов связывается с гемоглобином, отдавшим кислород, а накапливающиеся анионы НСО3 выходят из эритроцита в плазму :
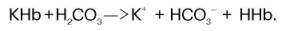
В обмен на эти ионы в эритроцит поступают анионы хлора, для которых мембрана эритроцитов проницаема, в то время как натрий – другой составной элемент хлорида натрия, содержащегося в крови, остается в плазме. В итоге в плазме крови повышается содержание бикарбоната натрия NaHCO3.
Этот процесс способствует восстановлению щелочного резерва крови, т.е. бикарбонатная буферная система находится в довольно тесных функциональных связях с буферной системой эритроцитов.
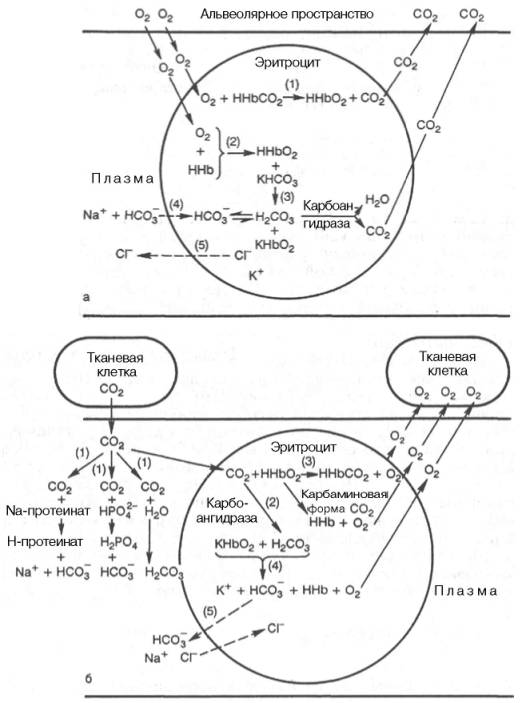
Рис. 17.7. Роль системы плазма-эритроцит в дыхательной функции крови (по
Г.Е. Владимирову, Н.С. Пантелеевой).
а — химические процессы в капиллярах легких; б — химические процессы в капиллярах ткани.
Образующаяся угольная кислота быстро расщепляется при участии карбоангидразы на углекислый газ и воду. Низкое РCO2 в просвете альвеол способствует диффузии углекислого газа из эритроцитов в легкие.
По мере снижения в эритроцитах концентрации бикарбоната из плазмы крови в них поступают новые порции ионов НСО3 – , а в плазму выходит эквивалентное количество ионов Сl – . Концентрация бикарбоната натрия в плазме крови в легочных капиллярах быстро падает, но одновременно в плазме повышается концентрация хлорида натрия, а в эритроцитах свободный гемоглобин превращается в калийную соль оксигемо-глобина.
Итак, в форме бикарбоната при участии гемоглобина эритроцитов транспортируется с кровью к легким более 80% от всего количества углекислого газа.
Механизмы переноса углекислого газа от тканей кровью
В сложных механизмах транспорта газов кровью и газообмена в тканях важная роль отводится эритроцитам, ответственным за доставку О2 к различным органам и удаление образующегося в процессе метаболизма СО2.
Эритроцит – безъядерная клетка, лишенная митохондрий, основным источником энергии для эритроцита служит глюкоза, метаболизируемая в гексозомонофосфатном шунте или цикле Эмбдена-Мейергофа. Транспорт О2 обеспечивается в значительной мере гемоглобином, состоящим из белка глобина и гема. Последний представляет собой комплексное соединение железа и порфирина. Глобин представляет собой тетрамер полипептидной цепи. Hb A (HbA) – основной гемоглобин взрослых содержит 2 – альфа и 2 – бета – цепи, Hb A2 – содержит две альфа и две дельта цепи.
Читайте также: Вязание комбинируем с тканью для женщин
Гем состоит из иона железа, встроенного в порфириновое кольцо. Ион железа гема обратимо связывает одну молекулу О2. С одной молекулой Hb максимально связываются 4 молекулы О2 с образованием оксигемоглобина.
Гем может подвергаться не только оксигенации, но и истинному окислению, когда железо становится из двухвалентного трехвалентным. Окисленный гем носит название гематина, а молекула гемоглобина становится метгемоглобином. В крови человека метгемоглобин находится в незначительных количествах, его уровень резко возрастает при отравлениях. Метгемоглобин не способен отдавать кислород тканям.
В норме метгемоглобин составляет менее 3% общего Hb крови. Основная форма транспорта О2 – в виде оксигемоглобина. Кислород транспортируется артериальной кровью не только в связи с гемоглобином, но и в растворенном виде. Принимая во внимание тот факт, что 1 г Hb может связать 1,34 мл О2, кислородная емкость крови в среднем у взрослого человека составляет около 200 мл/л крови. Одним из показателей кислородного транспорта является насыщение артериальной крови О2(Sa O2), равного отношению О2, связанного с Hb, к кислородной емкости крови:
SaO2=O2, связанного с Hb/O2 емкость крови* 100%.
В соответствии с кривой диссоциации оксигемоглобина насыщение артериальной крови кислородом в среднем составляет 97%, в венозной крови – 75%.
PaO2 в артериальной крови около 100 мм. рт. ст., а в венозной – около 40 мм. рт. ст.
Количество растворенного кислорода в крови пропорционально парциальному давлению О2 и коэффициэнту его растворимости.
Последний для О2 составляет 0,0031/100 мл крови/ 1 мм. рт. ст.. Таким образом, 100 мл крови при PaO2, равном 100 мм. рт. ст., содержит менее 0,31 мл O2.
Диссоциация оксигемоглобина в тканях обусловлена главным образом химическими свойствами гемоглобина, а также рядом других факторов – температурой тела, рН среды, р СО2.
При понижении температуры тела наклон кривой диссоциации оксигемоглобина возрастает, а при ее повышении – снижается, и соответственно снижается сродство Hb к О2.
При снижении рН, т.е. при закислении среды, сродство гемоглобина к О2 уменьшается. Увеличение напряжения в крови СО2 также сопровождается снижением сродства Hb к О2 и уплощением кривой диссоциации оксигемоглобина.
Известно, что степень диссоциации оксигемоглобина определяется содержанием в эритроцитах некоторых фосфорорганических соединений, главным из которых является 2,3 – ДФГ (2,3 дифосфоглицерат), а также содержанием в эритроцитах катионов. В случаях развития алкалозов, поглощение О2 в легких увеличивается, но в то же время затрудняется отдача кислорода тканями. При ацидозах наблюдается обратная картина.
4.2.Утилизация кислорода тканями
Тканевое или клеточное дыхание включает три стадии. На первой стадии пируват, аминокислоты и жирные кислоты окисляются до двухуглеродных фрагментов ацетильных групп, входящих в состав ацетилкофермента А. Последние на втором этапе окисления включаются в цикл лимонной кислоты, где происходит образование высокоэнергетических атомов водорода и высвобождение СО2 – конечного продукта окисления органических субстратов. На третьей стадии клеточного дыхания атомы водорода делятся на протоны (Н+) и «высокоэнергетические» электроны, передающиеся по дыхательной цепи на молекулярный О2 и восстанавливающие его до НО2. Перенос электронов сопряжен с запасом энергии в форме АТФ, т.е. с окислительным фосфорилированием (рис.6).
Касаясь патогенеза метаболических сдвигов, свойственных гипоксическим состояниям, следует отметить, что в организме человека более 90% всего потребляемого кислорода восстанавливается с участием цитохромоксидазы митохондрий, и лишь около 10% кислорода метаболизируется в тканях с участием оксигеназ: диоксигеназы и монооксигеназы.
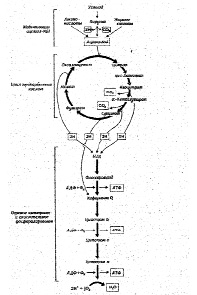
Рис.6. Схема тканевого дыхания. Конечные продукты каждой стадии даны в рамке (Ленинджер А., 1999)
Наиболее многочисленны и сложны монооксигеназные реакции, протекающие в эндоплазматическом ретикулуме клеток при участии цитохрома Р-450 и обеспечивающие гидроксилирование субстрата (стероидных гормонов, лекарственных препаратов и различных др. соединений) и, как правило, его инактивацию.
Диоксигеназы катализируют реакции, в которых в молекулу органического субстрата включаются оба атома молекулы кислорода (например, реакция окисления катехола молекулярным кислородом с раскрытием кольца).
В реакциях, связанных с переносом электронов, т.е. в реакциях окисления-восстановления, где, как указывалось выше, используется более 90% потребляемого кислорода, атомы водорода, отщепленные дегидрогеназами от субстратов в цикле лимонной кислоты, передают свои электроны в цепь переноса электронов и превращаются также в Н +. Как известно, помимо 4 пар атомов водорода, поставляемых каждым оборотом цикла лимонной кислоты, образуются и другие атомы водорода, отщепленные дегидрогеназами от пирувата, жирных кислот и аминокислот в процессе их расщепления до Ацетил-СоА и других продуктов.
Читайте также: Как проявляется активность ферментов в живых тканях
Таким образом, все атомы водорода, отщепляемые дегидрогеназами от субстратов, передают свои электроны в дыхательную цепь к конечному акцептору электронов – кислороду.
Касаясь последовательности транспорта электронов в окислительно-восстановительных реакциях, протекающих на внутренней мембране митохондрий, прежде всего, следует отметить, что от всех НАД – зависимых реакций дегидрирования восстановленные эквиваленты переходят к митохондриальной НАДН – дегидрогеназе, затем через ряд железосерных ферментов передаются на убихинон М цитохрому b. Далее электроны переходят последовательно на цитохромы С1 и С, затем на цитохромы аа 3 (цитохромоксидазу – медьсодержащий фермент). В свою очередь цитохромоксидаза передает электроны на кислород. Для того, чтобы полностью восстановить кислород с образованием 2-х молекул воды требуется 4 электрона и четыре Н+ .
Скорость утилизации О2 в различных тканях различна. В среднем взрослый человек потребляет 250 мл О2 в 1 мин. Максимальное извлечение О2 из притекающей артериальной крови свойственно миокарду.
Кислород используется в клетках, в основном в метаболизме белков, жиров, углеводов, ксенобиотиков, в окислительно-восстановительных реакциях в различных субклеточных фракциях: в митохондриях, в эндоплазматическом ретикулуме, в реакциях липопероксидации, а также в межклеточном матриксе и в биологических жидкостях.
Коэффициент утилизации О2 в тканях равен отношению потребления О2 к интенсивности его доставки, широко варьирует в различных органах и тканях.
В условиях нормы минимальную потребность в О2 проявляют почки и селезенка, а максимальную потребность – кора головного мозга, миокард и скелетные мышцы, где коэффициент утилизации О2 колеблется от 0,4 до 0,6, а в миокарде до 0,7. При крайне интенсивной физической работе коэффициент утилизации О2 мышцами и миокардом может возрастать до 0,9.
Обмен дыхательных газов в тканях происходит в процессе свободной и облегченной диффузии. При этом О2 переносится по градиенту напряжения газа из эритроцитов и плазмы крови в окружающие ткани.
Одновременно происходит диффузия СО2 из тканей в кровь. На выход О2 из крови в ткани влияет диссоциация оксигемоглобина в эритроцитах, что обеспечивает так называемую облегченную диффузию О2. Интенсивность диффузионного потока О2 и СО2 определяется градиентом их напряжения между кровью и тканями, а также площадью газообмена, плотностью капилляров, распределением кровотока в микроциркуляторном русле. Интенсивность окислительных процессов в тканях определяется величиной критического напряжения О2 в митохондриях, которое в условиях нормы должно превосходить 0,1-1 мм рт. ст.
Соответствие доставки О2 к органам и тканям, возросшим потребностям в оксигенации обеспечивается на клеточном, органном уровнях за счет образования метаболитов изнашивания, а также при участии нервных, гормональных и гуморальных влияний.
Основная масса углекислого газа (СО2) образуется в организме как конечный продукт различных метаболических реакций и транспортируется к легким с кровью. Вдыхаемый воздух содержит лишь незначительное количество СО2.
Транспорт СО2 кровью осуществляется в 3-х состояниях: в виде аниона бикарбоната, в растворенной форме и в виде карбаминовых соединений.
СО2 хорошо растворяется в плазме крови и в артериальной крови, около 5% от общей двуокиси углерода содержится в крови в растворенной форме.
Анион бикарбоната составляет около 90% от общего содержания СО2 в артериальной крови: СО2+Н2О — Н++НСО-3.
Эта реакция медленно протекает в плазме крови, но чрезвычайно интенсивно происходит в эритроцитах при участии фермента карбоангидразы. Мембрана эритроцита относительно непроницаема для Н+, как и вообще для катионов, но в тоже время, проницаема для ионов НСО-3, выход которых из эритроцитов в плазму обеспечивается притоком Cl- из плазмы в эритроциты. Часть Н+ забуферивается гемоглобином с образованием восстановленного гемоглобина.
Третьей формой транспорта СО2 кровью являются карбаминовые соединения, образованные взаимодействием СО2 с концевыми группами белков крови преимущественно с гемоглобином:
Hb NH2 + CO2 — Hb NH COOH > Hb NH COO + Н+
Карбаминовые соединения составляют около 5% от общего количества СО2, транспортируемого кровью.
В оксигенированной артериальной крови напряжение СО2 составляет 40 мм. рт. ст., а в венозной крови Рv СО2 равно 46 мм. рт. ст.
4.4.Связывание гемоглобина с окисью углерода
Угарный газ (СО) – окись углерода обладает значительно большим сродством к гемоглобину, чем О2, с последующим образованием карбоксигемоглобина. СО входит в состав бытового газа, а также выделяется при работе двигателей внутреннего сгорания. При концентрации СО во вдыхаемом воздухе всего в количестве 7*10- 4 около 50% гемоглобина превращается в карбоксигемоглобин. В норме в крови содержится около 1% HbCO, у курильщиков – 3% . В крови водителей такси концентрация карбоксигемоглобина достигает 20%. Карбоксигемоглобин диссоциирует с отдачей О2 в 200 раз медленней оксигемоглобина и в то же время препятствует его диссоциации в тканях.
- Правообладателям
- Политика конфиденциальности