
Сопротивление нормальных водорослей почти в 10 раз больше сопротивления морской воды, тогда как мертвая ткань имеет сопротивление почти равное сопротивлению морской воды. Вообще Са понижает проницаемость, а Mg, Na и К ее увеличивают. [18]
Следовательно, основное различие влияния низких температур на живую и мертвую клетки заключается в том, что в мертвой ткани максимальное накопление льда происходит при более высокой температуре, чем в ткани живого организма, в которой кристаллообразование протекает с некоторым запаздыванием. [19]
Особенность паразитов такого типа — способ заселения тканей растения, а именно сначала они должны убить живые клетки растения своими токсичными выделениями, а после этога использовать мертвые ткани для питания. [20]
В связи с этим огромное значение приобретают результаты исследований [13], в которых установлено, что в тканях с интенсивным обменом концентрация свободных радикалов ( оцененная методом электронного магнитного резонанса) выше, чем в мертвых тканях или тканях в состоянии покоя. Приведенные результаты подтверждают представление о том, что в живых организмах под влиянием кислорода действительно происходят синглетно-триплетные переходы. [21]
Бедфорда была доставлена на самолете в хранилище в Аризоне, где она будет находиться вплоть до того времени, когда найдут способ лечения рака в после /: — ней стадии, а заодно и способ оживления мертвых тканей . [22]
Начало кристаллообразования в межклеточном пространстве происходит, как правило, только в случае замораживания живой ткани — живой рыбы, свежих плодов и овощей. В мертвой ткани кристаллизационный центр может возникнуть как в межклеточном пространстве, так и внутри клетки или же одновременно и в клетке и в межклеточном пространстве. [23]
Кривая 2 соответствует мертвой ткани. В мертвой ткани разрушены мембраны, и она не обладает емкостным сопротивлением, поэтому нет частотной зависимости импеданса. [24]
Начало кристаллообразования в межклеточном пространстве происходит, как правило, только в случае замораживания живой ткани — живой рыбы, свежих плодов и овощей. В мертвой ткани кристаллизационный центр может возникнуть как в межклеточном пространстве, так и внутри клетки или же одновременно и в клетке и в межклеточном пространстве. [25]
Бактерицидное действие сохраняется 18 — 24 час. Препарат растворяет мертвые ткани , но не разрушает, как иод, живые. В хлоркозане можно растворить до 8 5 — 10 % дихлорамида, причем растворы его являются относительно стойкими. [26]
В мертвом теле насекомого-хозяина продолжается развитие нематод. Вероятно, мертвые ткани служат источником питания, а находящийся в них резервуар патогенных бактерий мигрирующие личинки покоящейся стадии переносят дальше. [27]
Совершенно иная картина наблюдается при замораживании мертвой ткани. Криоскопическая температура соков мертвой ткани лежит значительно выше криоскопической температуры ткани живого организма. [28]
Во флоэме заметны сезонные изменения размеров клеток, аналогичные образованию ранней и поздней древесины ( см. рис. 9.1) и связанные со старением, подобно образованию ядровой древесины. Корка в основном представляет мертвую ткань , в клетках которой появляются различные отложения. [29]
Многие органические конечные продукты метаболизма откладываются у растений в омертвевших постоянных тканях ( таких, как ядровая древесина), а также в листьях или коре, которые периодически сбрасываются. Многолетние растения состоят в основном из мертвых тканей . Экскреты скапливаются в этих тканях и уже не могут оказывать вредного воздействия на активность живых тканей. Аналогичным образом могут накапливаться многие минеральные соли, поглощаемые растением в виде ионов. Некоторые органические кислоты, вредные для растения, часто связываются с избыточными катионами и выпадают в виде безопасных нерастворимых кристаллов, которые могут храниться в клетках растения. Например, ионы кальция и сульфат-ионы поглощаются растением одновременно, но сульфат-ионы сразу же используются для синтеза аминокислот, а кальций остается в избытке. Ионы Са2 легко реагируют со щавелевой и пектовой кислотами, образуя с ними безвредные нерастворимые продукты — оксалат и пектат кальция. [30]
Читайте также: Полиэстер ткань для зонтов в
Виды эквивалентных электрических схем тканей организма
Электрические свойства биологических тканей моделируются сочетанием элементов, обладающих омическими и емкостными свойствами. Эти свойства можно моделировать с помощью эквивалентных электрических схем. Рассмотрим три из них, рис. 17.
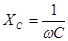
Схема I не совсем удовлетворительна, т.к. содержит бесконечно большое сопротивление постоянному току (емкость не проводит постоянный ток), чего не наблюдается в тканях организма. Поэтому при низкой частоте схема I даст существенные отклонения расчетных значений импеданса (полного сопротивления) от реального импеданса биоткани. В схеме // при увеличении частоты тока ω емкостное сопротивление стремится к нулю поэтому импеданс схемы также стремится к пулю. Это противоречит опыту.
У живых объектов импеданс уменьшается по мере увеличения частоты только до определенного значения. Для живых тканей характерно более сложное сочетание параллельного и последовательного соединений элементов, например, схема ///.
В состоянии покоя реактивное сопротивление ткани носит только емкостный характер, при возбуждении ткань приобретает индуктивные свойства и обладает индуктивным сопротивлением
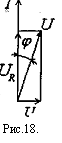
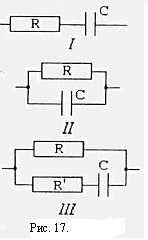
При анализе электрических свойств ткани рассматриваем только состояние покоя. Найдем импеданс Z простейшей эквивалентной электрической схемы /, рис. Векторная диаграмма имеет вид, показанный на рис.17. Общее напряжение равно
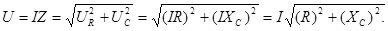
Следовательно, импеданс цепи равен
Угол сдвига фазы между током I и напряжением U найдем из условия.
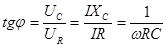
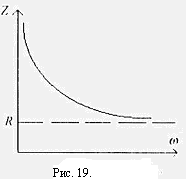 На рис.19. показана зависимость Z импеданса эквивалентной электрической схемы / (рис.17.) от частоты, построенная но формуле
На рис.19. показана зависимость Z импеданса эквивалентной электрической схемы / (рис.17.) от частоты, построенная но формуле 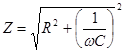
Как видно из рис. 19., импеданс цепи монотонно падает с увеличением частоты электрического тока, постепенно приближаясь к активному сопротивлению R..
Величины логарифмов импеданса и относительной диэлектрической проницаемости можно считать по модулю примерно пропорциональными друг другу.
Зависимость импеданса (или относительной диэлектрической проницаемости) биологических тканей от частоты электрического тока называется дисперсией импеданса (или относительной диэлектрической проницаемости), а график этой зависимости дисперсионной кривой.
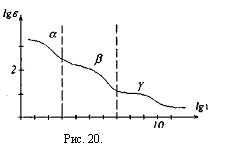
В отличие от зависимости импеданса от частоты электрического тока для эквивалентной электрической схемы биологической ткани, реальная дисперсионная кривая, например, скелетной мышцы, немонотонно падает с увеличением частоты, рис.20.
При действии на ткани организма переменного электрического тока или переменного электрического поля наблюдаются три области дисперсии. Область а — дисперсии.
Наблюдается до частот 1кГц (lgv≈3)
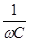
В низкочастотной части спектра области а- дисперсии электрический ток течет практически только по межклеточной жидкости, т.к. сопротивление мембран клеток очень велико. В высокочастотной части спектра импеданс падает за счет снижения емкостного сопротивления мембран клеток Хс = Возникает ток смещения, текущий через мембрану.
Область β- дисперсии.Наблюдается до частот I0 4 кГц (Igv≈3 – 7)
За счет увеличения частоты переполяризации мембран клеток снижается емкостное сопротивление ткани. Переполяризация рассматривается относительно пулевого уровня потенциала па мембране, за который принят потенциал покоя.
Читайте также: Дымчатый кварц цвет ткани
Затраты энергии на поляризацию и нереполяризацию клеточных структур (мембран, белковых молекул, молекул воды, атомов и т.д.) определяют импеданс ткани. Снижение этих затрат приводит к снижению импеданса биоткани.
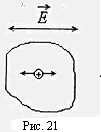 Многие ионы внутри клеток, рис.21, при такой частоте колебаний напряженности электрического поля
Многие ионы внутри клеток, рис.21, при такой частоте колебаний напряженности электрического поля 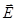 не успевают дойти до клеточных и внутриклеточных мембран. Для них сопротивление мембран роли не играет.
не успевают дойти до клеточных и внутриклеточных мембран. Для них сопротивление мембран роли не играет.
В высокочастотной области β- дисперсии существенное значение приобретает структурная поляризация и переполяризация крупных, в частности белковых молекул. Структурная поляризация заключается в том, что внутри молекулы, под действием электрического поля, смещаются ионы и ионные группы, входящие в состав молекул, что влияет на функциональные свойства белков.
Область γ дисперсии. Наблюдается до частот 10 7 кГц (Igv =• 7 — 10).
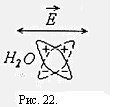
За счет высокой частоты колебаний электрического тока, емкостное сопротивление мембран очень мало и они не представляют большого препятствия для переменного тока. Поэтому сопротивление биоткани в общем мало.
В низкочастотной части спектра области γ- дисперсии уменьшение диэлектрической проницаемости обусловлено постепенным ослаблением ориентационной переполяризации дипольных молекул воды, рис.22. В этой области важный вклад в импеданс ткани вносит структурная поляризация крупных молекул. Энергия электрического тока с увеличением частоты все меньше теряется на переориентацию молекул воды и структурную переполяризацию крупных молекул, поэтому уменьшается импеданс ткани.
В высокочастотной части спектра γ — дисперсии молекулы воды под действием переменной напряженности Е почти не переполяризуются. Уровень электрического сопротивления определяется электронной переполяризацией атомов, т. с. возникновением переменных атомных диполей, рис.23.
МЕТОД ЭЛЕКТРОПРОВОДНОСТИ В МЕДИЦИНЕ
ИМПЕДАНС БИОЛОГИЧЕСКИХ ТКАНЕЙ
ПРИРОДА ЕМКОСТНЫХ СВОЙСТВ ТКАНЕЙ ОРГАНИЗМА
Как показывает опыт, электропроводность растворов не зависит от частоты переменного тока. При изучении электропроводности биологических объектов обнаружили, что их сопротивление на высоких частотах (
10 7 Гц) гораздо меньше, чем на низких. На рис.9 (кривая 1) представлена зависимость сопротивления мышечной ткани от частоты (кривая дисперсии). Дисперсия электропроводности обычно наблюдается в интервале частот 10 2 ÷ 10 8 Гц. Наличие дисперсии импеданса для живых тканей обусловлено тем, что при низких частотах (как и для постоянного тока) на величину электропроводности значительное влияние оказывает макроструктурная поляризация в тканях. По мере увеличения частоты поляризационные явления сказываются всё меньше и меньше.
Отмирание ткани ведёт к росту проницаемости мембран, при этом крутизна дисперсии уменьшается (рис.9, кривая 2). Для мёртвой ткани поляризация на границах раздела практически полностью исчезает и явление дисперсии импеданса больше не наблюдается (рис.9, кривая 3). Т.о. крутизна графика зависимости Z = f(ω) позволяет судить о жизнеспособности той или иной ткани.
Наличие дисперсии электропроводности говорит о том, что в биологических объектах наряду со структурами, которые оказывают переменному току активное сопротивление, есть и такие, которые обладают реактивным сопротивлением. Как оказалось, структуры с индуктивным сопротивлением (подобные катушкам) в биологических организмах отсутствуют.
Клеточные мембраны, омываемые с одной стороны тканевой жидкостью, а с другой цитоплазмой, представляют собой системы подобные конденсатору. В тканях имеются так же макроскопические образования, состоящие из различных непроводящих соединительных оболочек и перегородок, по обе стороны которых находятся ткани, хорошо проводящие электрический ток. Это так же придаёт тканям емкостные свойства.
В целом, сопротивление биологических тканей будет определяться суммой омического и емкостного сопротивления:
Присутствие в биологических организмах структур с ёмкостным сопротивлением подтверждается также наличием сдвига фаз между током и напряжением. Для биологических систем характерна большая величина этого угла, например, на частоте 1000 Гц: кожа человека – φ = 55 0 , мышца кролика – φ = 65 0 , нерв лягушки – φ = 64 0 . Это показывает, что доля емкостного сопротивления в импедансе тканей велика.
Читайте также: Из чего делают рогожку рогожка ткань
При моделировании электропроводности живых клеток и тканей прибегают к эквивалентным схемам, т.е. к таким комбинациям омического сопротивления и ёмкости, которые в некотором приближении могут отражать характер течения тока и значения электрических параметров клетки и тканей. Простейшими из таких моделей являются схемы с последовательным и с параллельным соединением R и C – (рис.10а и 11а). Но эти простейшие схемы не отражают истинного положения дел, т.к. графики зависимости Z от ω противоречат данным по живым тканям (сравните рис.10б, 11б и рис.6, кривая1).
Наиболее удачной моделью является схема, представленная на рис.12а. На этой схеме Rк,1 и Rк,2 – активные сопротивления кожи на входе и выходе тока; Rт – общее омическое сопротивление подкожных тканей; C1, C2 и C3 – конденсаторы, моделирующие биологические структуры, обладающие ёмкостным сопротивлением. Стрелками показан маршрут переменного тока, т.е. показаны структуры, через которые проходит ток на каждом из участков биологического организма между электродами за один полупериод. Зависимость сопротивления этой эквивалентной схемы от частоты хорошо согласуется с кривой дисперсии импеданса для биотканей (рис.12б) Существуют и другие эквивалентные схемы, однако ни одна из них в точности не может воспроизвести закономерности течения переменного тока, присущие биологическим системам.
Измерение электропроводности биологических тканей для переменного тока широко используется в диагностике, а так же в биологических и медицинских исследованиях. Например, значительное возрастанию импеданса ткани на низких частотах позволяет обнаружить воспаление уже на первых стадиях. Некоторые заболевания щитовидной железы диагностируются по изменению угла сдвига фаз между током и напряжением. Для характеристики физиологического состояния тканей используют также величину крутизны кривой дисперсии. Этот критерий применяют, например, при оценке жизнеспособности ткани, предназначенной для трансплантации.
Импеданс тканей существенным образом зависит от кровенаполнения сосудов. Кровь имеет меньшее сопротивление, чем стенки сосудов или клетки, поэтому во время систолы полное сопротивление ткани уменьшается, а при диастоле – увеличивается. Диагностический метод, основанный на регистрации импеданса тканей в процессе сердечной деятельности, называется реографией (импеданс – плетизмографией). С помощью этого метода получают реограммы головного мозга (реоэнцефалограмма), сердца (реокардиограмма), лёгких, печени, сосудов, конечностей. Измерения обычно проводят на частоте 30 кГц.
Переменным называется ток, изменяющийся с течением времени по величине и направлению. Потребляемый, промышленный ток является синусоидальным. Мгновенное значение его параметров меняются со временем по закону синуса (или косинуса):
Переменный ток характеризуется периодом Т, частотой ν = 1/Т, циклической частотой , фазой φ = (ωt + φ0) Графически значения напряжения и силы переменного тока на участке цепи будут представляться двумя синусоидами, в общем случае сдвинутыми по фазе.
Для характеристики переменного тока вводится понятие действующего (эффективного) значения тока. Эффективным значением силы переменного тока называется сила такого постоянного тока, который выделяет в данном проводнике столько же тепла за время одного периода, сколько выделяет тепла и данный переменный ток.
Приборы, включенные в цепь переменного тока (амперметр, вольтметр) показывают эффективные значения тока и напряжения.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом