
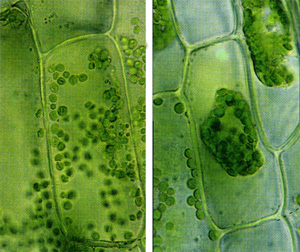
Группы клеток, сходные по строению, происхождению и выполняемым функциям, образуют ткани.
Из тканей построены органы и системы органов. Разные органы растений вместе образуют единый организм:
группа клеток —> ткань —> орган —> организм
У растений различают 6 видов тканей: образовательную, покровную, основную, опорную, проводящую и выделительную.
1.Образовательная ткань
Образовательная ткань находится на верхушке побега и на верхушке корня. Ее клетки плотно прилегают друг к другу. У них тонкие оболочки. За счет деления клеток растения растут.
Рост побега в длину и разрастание листьев, утолщение стеблей и корней, восстановление поврежденных мест деревьев — функции образовательной ткани. Из клеток образовательной ткани образуются все другие виды тканей.
Со временем клетки утрачивают способность делиться. Они становятся клетками постоянных тканей, таких как покровные, основные, проводящие и др.

2.Покровная ткань
Покровная ткань формируется на поверхности органов. Она представлена кожицей, пробкой и коркой. Защищает растения от высыхания, солнечных ожогов, неблагоприятных условий внешней среды.
Клетки кожицы — эпидермис — образуются на всех молодых органах растений. Эпидермис обеспечивает газообмен, испарение, всасывание, предохраняет органы растений от высыхания.
Но для зимующих растений это ненадежная защита. Вместо него перед наступлением зимы образуется пробка. Эта многослойная ткань состоит из мертвых, плотно прилегающих друг к другу клеток. Она защищает растения.
Корка — это наружная часть коры. Как и пробка, она состоит из мертвых клеток и защищает стволы и ветви от излишнего испарения, перегрева, вымерзания, ожога солнечными лучами, объедания животными.

3.Основная ткань
Основная ткань состоит из живых клеток и образует основу всех органов растения.
В зависимости от функции она подразделяется на фотосинтезирующую и запасающую.
Клетки фотосинтезирующей ткани содержат хлоропласты. В них осуществляется фотосинтез. Основная масса этой ткани сосредоточена в листьях, меньшая часть — в молодых зеленых стеблях.
Запасающая ткань плодов, семян, стеблей, луковиц, листьев, корнеплодов, корневищ участвует в накоплении питательных веществ, которые необходимы прежде всего многолетним растениям.
Часть клеток основной ткани служит для запасания воды. Водоносная ткань содержится в основном в стеблях и листьях растений пустынных мест обитания и солончаков, например в стеблях кактусов или листьях алоэ.
Воздухоносная ткань рыхлая. У нее хорошо развиты межклеточные пространства (межклетники), в которые проникает воздух. Особенно хорошо они сформированы у растений, произрастающих в воде (водные и болотные) и на глинистой почве.
По воздухоносным межклетникам кислород доставляется к тем частям растения, связь которых с атмосферой затруднена.

Основная ткань (фотосинтезирующая и запасающая)
4.Опорная или механическая ткань
Опорная, или механическая, ткань выполняет у растений функцию каркаса, опоры Она находится в стеблях, листьях и плодах растений. Опорная ткань придает упругость и прочность всем органам растений.
Поэтому при сильном ветре не ломаются хрупкие стебли, не разрываются большие листовые пластинки и листья не срываются с деревьев.

Опорная (механическая) ткань
В мякоти плодов груши, айвы, рябины, в семенах пальмы, в косточках вишни, сливы, абрикоса, персика встречаются каменистые клетки. Они тоже являются опорной тканью.
В органах молодых растений опорная ткань развивается не сразу. Например, косточки незрелых фруктов — сливы, вишни, абрикоса — мягкие, беловатого цвета. По мере созревания плодов их оболочка темнеет и становится твердой.
Семена от повреждений защищает опорная ткань, состоящая сначала из живых клеток. Позже они теряют цитоплазму, стенки утолщаются и древеснеют.
В размещении механической ткани в растительных органах существует особая закономерность. Изучая ее, человек учится у растений создавать прочные, экономичные, радующие глаз здания, башни, мосты, которые к тому же будут естественно вписываться в окружающую среду.
5.Проводящая ткань
Функции проводящей ткани заключаются в проведении воды и питательных веществ из одного органа растения в другой. Она состоит из двух частей.
Одна часть — ксилема, или древесина, — обеспечивает восходящий поток и доставляет воду и минеральные соли от корней в надземную часть растения.
Клетки древесины представляют собой полые трубки (сосуды) с одеревеневшими мертвыми стенками. В сосудах имеются отверстия, через которые вдоль всего сосуда осуществляется движение жидкости.
Другая часть — флоэма, или луб, — обеспечивает нисходящий поток, т. е. проведение образовавшихся в листьях органических веществ в подземные органы. В состав луба входят ситовидные трубки и клетки-спутницы. Луб и древесина расположены в стебле, корне, жилках листьев.

Проводящие ткани: Ксилема и Флоэма.
Читайте также: Заглаженная ткань как исправить
Органические вещества, образованные в листьях, доставляются к стеблям, корням, точкам роста, плодам, семенам по ситовидным трубкам. Клетки ситовидных трубок живые.
В поперечных перегородках члеников ситовидных трубок имеется большое количество мелких отверстий, как в сите.
У растений элементы проводящей, опорной и запасающей тканей образуют проводящие, или сосудисто-волокнистые, пучки. Они хорошо видны в листьях в виде жилок, распространены в стебле, корнях и плодах.

Осенью отверстия перегородок ситовидных трубок затягиваются мозолистым веществом, и ток органических веществ по трубке прекращается. Растение впадает в состояние покоя.
Весной мозолистое вещество растворяется, и ток по ситовидным трубкам возобновляется. Проводящая ткань осуществляет связь между корнем и побегом.
6.Выделительная ткань
Известно, что у растений нет специальных выделительных органов, как у животных. Но выделительные ткани есть у большинства растений. Ими представлены смоляные и эфирно-масляные ходы, железы, железистые волоски, нектарники и т. д.
Растения выделяют ароматические и сахаристые вещества, привлекающие насекомых-опылителей. Эфирные масла защищают растения от поедания травоядными животными.

Строение растительной клетки
Увеличительные приборы
Группы клеток, сходные по строению, происхождению и выполняемым функциям, образуют ткани. Из тканей построены органы и системы органов. Разные органы растений вместе образуют единый организм. Рост побега в длину и разрастание листьев, утолщение стеблей и корней, восстановление поврежденных мест деревьев функции образовательной ткани. Из клеток образовательной ткани образуются все другие виды тканей. Покровная ткань защищает растения от высыхания, солнечных ожогов, неблагоприятных условий внешней среды. Основная ткань состоит из живых клеток и образует основу всех органов растения. Опорная, или механическая, ткань выполняет у растений функцию каркаса, опоры.
Функции проводящей ткани заключаются в проведении воды и питательных веществ из одного органа растения в другой. У растений нет специальных выделительных органов. Но выделительные ткани есть у большинства растений. Ими представлены смоляные и эфирно-масляные ходы, железы, железистые волоски.
Биологический русско-английский глоссарий
Структурная организация фотосинтетического аппарата, строение листа как органа фотосинтеза, ультраструктура и онтогенез хлоропластов
Ответы на вопросы по физиологии растений
Основные принципы действия регуляторных механизмов растительной клетки; компартментация каталитических систем и метаболических фондов протопласта как один из механизмов регуляции клеточного метаболизма.
Клетка – это мельчайшая единица жизни, характеризующаяся определенным типом обмена веществ, самостоятельным энергетическим циклом и способностью к саморегуляции и саморазвитию на основе взаимодействия различных своих компо-нентов и взаимодействия с внешними условиями. В живой клетке нет изолированных процессов. Основой этого взаимодействия служит непрерывный обмен продуктами реакций, образующимися в органоидах определенного вида и поступающими в цитоплазму и другие органоиды.
Сложное строение растительной клетки, дифференцированной на большое количество специализированных органоидов и структур в соответствии с выполняемыми ее функциями, требует и совершенных систем регуляции. Регуляция обеспечивает целостность и гомеостаз организма, т. е. сохранение постоянства параметров внутренней среды, а также создает условия для его развития (эпигенеза).
В ходе эволюции сначала возникли внутриклеточные системы регуляции. К ним относятся регуляция на уровне ферментов, генетическая и мембранная регуляции. Все эти системы регуляции тесно связаны между собой. Принципы действия регуляторных механизмов клетки:
– принцип взаимодействия компонентов цитоплазмы.
В основе всех форм внутриклеточной регуляции лежит единый первичный принцип – рецепторно-конформационный принцип. Белковая молекула – фермент, рецептор или регуляторный белок — «узнает» специфический для нее фактор и, взаимодействуя с ним, изменяет свою конфигурацию. В мультикомпонентных комплексах ферментов, генов и мембран конформационные изменения молекул-рецепторов кооперативно передаются на весь комплекс, влияя на его функциональную активность.
Важнейшими в регуляторных механизмах клетки является принцип компар-тментации, обуславливающий высокую степень временной и пространственной ор-ганизованности метаболизма, и взаимодействия компонентов цитоплазмы. Взаимо-действие обеспечивается наличием гиалоплазмы и эндоплазматической сети.
Компартменты – дифференцированные, специализированные участки, или отсеки в протоплазме, различающиеся по степени активности содержащихся в них химических соединений и систем, регулирующих их превращения. Явление компартментации обуславливают клеточные мембраны, которые выполняют функцию расчленения биохимических процессов, разделения их между различными компонентами протоплазмы и пространственного размещения в объеме клетки фондов метаболитов и ферментов.
В растительной клетке имеется три основных компартмента: свободное пространство (СП), цитоплазма и вакуоль. Компартментальны отдельные участки клетки и органелл, отдельные участки мембран, отдельные фонды метаболитов, ферментов и всех других морфологических и химических компонентов протопласта.
Характерная особенность компартментированных метаболитов и ферментов — их подвижность, что достигается включением метаболитов в везикулы и мелкие вакуоли и концентрацией их в трубочках эндоплазматического ретикулума и других трубчатых структурах. Это способствует, например, отводу ассимилятов из хлоропластов без их соприкосновения со стромой цитоплазмы. Эффективность действия механизмов фотофосфорилирования в хлоропластах определяется компартментами, обеспечивающими образование протонного градиента. Компартментальны в данном случае мембранные диски гран хлоропластов. Практически все основные пути метаболизма углеводов прочно ассоциированы с определенными участками плазматических мембран. В клетке существуют компартменты, содержащие системы гликолиза, цикла Кребса и пентозофосфатного шунта. Так все участки цикла Кребса локализованы в матриксе или во внутренней мембране митохондрий. Аналогичные закономерности установлены для процессов биосинтеза и превращения аминокислот, поглощения и выделения минеральных веществ, воды, регуляции действия имеющихся в клетке ферментов и т. п.
Читайте также: Ткань мебельная scandinavia ширина 140 см цвет бежевый
С появлением многоклеточных организмов развиваются и совершенствуются межклеточные системы регуляции. Они включают в себя по крайней мере трофическую, гормональную и электрофизиологическую системы. Такие взаимодействия сразу обнаруживаются при культивировании частей и органов растений в изолированном виде. Для поддержания жизни изолированных частей в инкубационную среду необходимо добавлять трофические и гормональные факторы, в норме поступающие из других органов целого растения.
Важным также является взаимная координация процессов, реакций, координация накопления и выделения веществ и энергии клеткой. Так, нуклеиновые кислоты играют роль хранителя и передатчика наследственной информации (ДНК) и посредника ее реализации (РНК). Белки несут функции рабочих механизмов – это ферменты, которые входят в состав клеточных структур и многих активных соединений. Основной механизм регуляции внутриклеточных процессов связан с влиянием на ферменты разнообразных факторов. Это может осуществляться на генетическом уровне, если определяется состав ферментов или количество того или иного фермента в клетке. Второй тип регуляции – влияние на сам фермент, в результате чего активность его может стимулироваться или ингибироваться. Может иметь место также структурный уровень регуляции – влияние на образование и сборку клеточных структур (мембран, хлоропластов, рибосом, митохондрий и т. д.). Конкретными регуляторами внутриклеточных процессов могут быть фитогормоны, которые вырабатываются клетками или окружающими их другими клетками, или продукты реакций по принципу обратной связи (аллостерический эффект). Регуляция может осуществляться также путем транспортировки в клетку продуктов метаболизма и их предшественников, ионов, изменения формы регулируемого фермента (конформаций).
Структурная организация фотосинтетического аппарата, строение листа как органа фотосинтеза, ультраструктура и онтогенез хлоропластов
«Зеленый лист, или, вернее, микроскопическое зеленое зерно хлорофилла является фокусом, точкой в мировом пространстве, в которую с одного конца притекает энергия солнца, а с другого берут начало все проявления жизни на земле. Растение — посредник между небом и землею. Оно истинный Прометей, похитивший огонь с неба» ( К. А. Тимирязев ).
Основой фотосинтетического аппарата служат:
− у высших зелёных растений внутриклеточные органеллы – хлоропласты;
− у большинства водорослей − специализированные внутриклеточные органеллы – хроматофоры;
− у фотосинтезирующих бактерий и синезелёных водорослей – тилакоиды (мембраны их содержат пигмент бактериохлорофилл или бактериовиридин, а также др. компоненты реакций фотосинтеза), погруженные в периферические слои цитоплазмы.
Хлоропласты занимают 20–30% объёма растительной клетки. У водорослей, например хламидомонады, имеется один хлоропласт, в клетке высших растений содержится от 10 до 70 хлоропластов и более.
Хлорофилл и каротиноиды, погруженные в тилакоидные мембраны хлоропластов, собраны в функциональные единицы – фотосистемы, каждая из которых содержит примерно 250 молекул пигментов. Фотосистема – совокупность ССК (светособирающего комплекса), фотохимического реакционного центра и переносчиков электрона.
ССК растений расположен в мембранах тилакоидов, у цианобактерий основная его часть вынесена за пределы мембран в прикреплённые к ним фикобилисомы — палочковидные полипептидно-пигментные комплексы, в которых находятся различные фикобилины: на периферии фикоэритрины, за ними фикоцианины и аллофикоцианины, последовательно передающие энергию на хлорофилл a реакционного центра.
Роль ССК состоит в том, чтобы собирать и передавать энергию квантов света на небольшое количество молекул (молекулы-ловушки) реакционных центров П680 и П700 которые и осуществляют фотохимические реакции.
Органом фотосинтеза у высших растений является лист. Кроме фотосинтеза в жизни растений лист выполняет функции газообмена, транспирации, синтеза ряда органических веществ, в том числе и фитогормонов.
Листья, будучи боковыми органами, как правило, имеют более или менее плоскую форму, что способствует созданию максимальной фотосинтезирующей поверхности.
Основной тканью листа является паренхимный комплекс клеток, несущий хлоропласты – мезофилл (греч. mesos – средний и филл) (рисунок 1). Остальные ткани листа обеспечивают работу мезофилла и поддерживают связь с окружающей средой. Покровная ткань (эпидермис) регулирует газообмен и транспирацию, защищает лист от внешних воздействий. Проводящие ткани осуществляют отток и приток веществ, поддерживают нормальное оводнение фотосинтезирующих клеток. Механические ткани совместно с живыми тургесцентными клетками мезофилла и эпидермиса образуют опорную систему листа.
Читайте также: Тип животной ткани у скелета
Мезофилл. Представлен клетками основной паренхимы, расположенными между верхним и нижним эпидермисом (исключая механические ткани и проводящие пучки). Состоит из живых клеток, с тонкими оболочками, округлой или слегка вытянутой формы, реже с небольшими выростами. Благодаря многочисленным хлоропластам мезофилл окрашен в зеленый цвет, так называемая хлоренхима листа. Иногда оболочки клетки образуют складки внутрь полости клетки, что увеличивает рабочую поверхность хлоренхимных клеток.
У типичных дорсовентральных листьев мезофилл неоднороден и диффе-ренцирован на столбчатую (палисадную) и губчатую паренхиму.
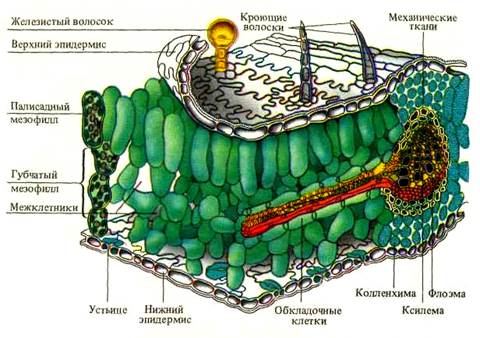
Рисунок 1 – схема строения листа
Клетки столбчатой паренхимы плотно сомкнуты, имеют продолговатую форму и располагаются перпендикулярно к верхней стороне листа. Столбчатый мезофилл может быть однослойный, иногда двуслойный, редко (у светолюбивых растений) многослойный. Основная функция палисадной паренхимы – ассимиляция углекислого газа (фотосинтез). Палисадная ткань располагается в наилучших условиях освещения, непосредственно под верхней эпидермой. Благодаря тому, что ее клетки вытянуты перпендикулярно к поверхности листа, у них имеется возможность регулировать направление и расположение хлоропластов таким образом, чтобы избежать повреждающего действия прямой солнечной радиации на фотосинтезирующий аппарат.
Губчатую паренхиму составляют относительно округлые клетки с большими межклетниками, которые занимают больший объем, чем сами клетки. Рыхлая структура мезофилла обусловливает большую общую площадь поверхности клеток, обращенной к межклетникам. Через крупные межклетники губчатого мезофилла идет газообмен.
Форма – овальные тельца длиной 5-10 мкм и диаметром 2-3 мкм. Количество в клетке листа – 15-20, у некоторых водорослей — 1-2 гигантских хлоропласта различной формы. Окраска – зеленая, обусловлена содержанием в хлоропластах хлорофилла.
Хлоропласты – двухмембранные органоиды (рисунок 2). Внутреннее содержимое хлоропластов − строма (матрикс) представляет собой гомогенную среду. Хлоропласты, как и другие органоиды, например, митохондрии, имеют ламеллярное строение, т.е. внутренние структуры органоида образованы инвагинациями внутренней мембраны – ламеллами. В хлоропластах внутренняя мембрана образует уплощенные инвагинации — тилакоиды, которые могут иметь форму дисков и в этом случае называются тилакоидами гран.
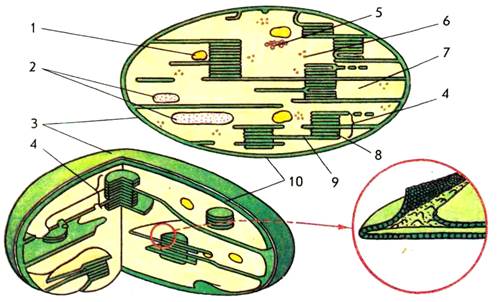
Рисунок 2 – схема строения хлоропласта:
1 – липидная капля; 2 – зерна крахмала; 3 – наружная мембрана; 4 – грана; 5 – ДНК; 6 – рибосома; 7 – строма; 8 – тилакоид граны; 9 – тилакоид стромы; 10 – внутренняя мембрана
Несколько лежащих друг над другом тилакоидов образуют стопку – грану. Другие тилакоиды, связывающие между собой граны или не контактирующие с ними, называются тилакоидами стромы. При таком строении значительно увеличивается фотоактивная поверхность хлоропластов и обеспечивается максимальное использование световой энергии. В мембранах тилакоидов локализованы зеленые (хлорофиллы), желтые и красные (каротиноиды) пигменты, компоненты редокс-цепей и запасания энергии, участвующие в поглощении и использовании энергии света.
Граны часто имеют форму цилиндров размером 2 мкм, в которых пигментно-липидные слои чередуются с белковыми слоями. Морфологически тилакоиды не гомогенны. На внутренней поверхности их мембран имеются специфические структурные образования, названные Парком квантосомами.
Хлоропласты – системы, способные к автономному синтезу белков. В них присутствуют низко- и высокомолекулярная РНК, специфическая кольцевая ДНК и ферменты, активирующие аминокислоты. Хлоропласты обладают собственными рибосомами.
1) осуществление фотосинтеза (основная функция);
2) в хлоропластах происходит вся сложная цепь процессов превращений первичных продуктов фотосинтеза (наращивание углеродной цепи, образование и распад полимерных форм углеводов и др.);
Биохимические системы синтеза и превращения углеводов функционируют в строме хлоропластов. В ней же может откладываться крахмал. В хлоропластах представлен весь набор биохимических систем, участвующих в синтезе АТФ.
Онтогенез хлоропластов. При нормальном освещении пропластиды пре-вращаются в хлоропласты. Сначала они растут, при этом происходит образование продольно расположенных мембранных складок от внутренней мембраны. Одни из них простираются по всей длине пластиды и формируют ламеллы стромы; другие образуют ламеллы тилакоидов, которые выстраиваются в виде стопки и образуют граны зрелых хлоропластов.
Несколько иначе развитие пластид происходит в темноте. У этиолированных проростков происходит в начале увеличение объема пластид – этиопластов, но система внутренних мембран не строит ламеллярные структуры, а образует массу мелких пузырьков, которые скапливаютсяя в отдельные зоны и даже могут формировать сложные решетчатые структуры (проламеллярные тела). В мембранах этиопластов содержится протохлорофилл, предшественник хлорофилла желтого цвета. Под действие света из этиопластов образуются хлоропласты, протохлорофилл превращается в хлорофилл, происходит синтез новых мембран, фотосинтетических ферментов и компонентов цепи переноса электронов.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом