
МЕТАПЛАЗИЯ (metaplasia; греч. metaplasso преобразовывать, превращать) — стойкое превращение одного типа ткани в другой, отличающийся от исходного морфологически и функционально. Метаплазия является приспособительным процессом, с помощью к-рого ткань выживает в изменившихся условиях существования благодаря структурной перестройке, в отличие от гистологической аккомодации, при к-рой клетки не изменяют тканеспецифических структурных свойств. Термин был впервые предложен в 1884 г. Р. Вирховом. Учение о Метаплазии интенсивно разрабатывалось в первой половине 20 в. в работах Ганземанна (D. P. Hansemann), Рибберта (М. W. H. Ribbert), Кромпехера (E. Krompecher), H. Г. Хлопина, Г. Э. Корицкого и др.
Различают прямую и непрямую Метаплазию. Прямая Метаплазия характеризуется изменением характера ткани без размножения ее клеток. Типичным примером прямой М. является превращение соединительной ткани в костную без участия остеогенных элементов. При непрямой, или неопластической, М. наблюдается пролиферация клеток с их размножением и дифференцировкой. Иногда новообразованная ткань может достигать более высокой степени развития, чем это было свойственно исходной ткани, напр, образование печеночной ткани при М. эпителия желчных протоков, образование панкреатических островков при М. эпителия протоков поджелудочной железы. Тогда процесс обозначают как прозопластическую М. В ряде случаев дифференцировка клеток оказывается более низкой, чем в исходной ткани, что называют регрессивной М., напр. при развитии многослойного плоского эпителия на месте цилиндрического мерцательного эпителия слизистой оболочки дыхательных путей.
Причиной, приводящей к М., являются чаще всего хрон, воспаление, дисгормональные изменения, опухолевый процесс, авитаминоз А.
В зависимости от локализации изменений выделяют М. эпителия, соединительной ткани с превращением ее в костную и хрящевую ткань и так наз. миелоидную М. с превращением лимфоидных клеток в клетки миелопоэтического ряда. Чаще встречается эпителиальная М., примером к-рой может служить превращение эпителия сальных и слюнных желез в условиях хрон, воспаления в многослойный плоский ороговевающий эпителий, эпителия молочных и потовых желез при дисгормональных изменениях — в бледный эпителий апокринного типа. Известна так наз. кишечная М., или энтеролизация слизистой оболочки желудка, напр, при гастрите, а также эпидермоидная М. эпителия желчного пузыря, желчных путей и протоков поджелудочной железы, возникающая при хрон, воспалении. Переходный эпителий мочевых путей при хрон, воспалении может подвергаться М. как в плоский многослойный эпителий, так и в железистый призматический. В мезодермальной эпителии отмечается возможность превращения мезотелия в фибробласты.
По мнению ряда авторов, М. может осуществляться в широких пределах и возможно превращение эпителия в соединительную ткань (см. Мезенхимопластика). Другие авторы ограничивают М. рамками гистогенетического типа, отрицая возможность превращения эпителия, напр, энтодермального типа, в эпидермальный. Нек-рые авторы отрицают возможность М., объясняя появление чуждого данному месту эпителия эмбриональной дистопией, гетероплазией, прозоплазией, плюрипотентностью камбиальных элементов и различными направлениями их дифференцировки. Наибольшее признание получила точка зрения, признающая возможность широких превращений эпителиальной ткани, но отрицающая вероятность превращения эпителия в соединительную ткань.
Клин, значение М. зависит от этиологии и локализации патол, процесса. В слизистой оболочке канала шейки матки и в эндометрии обнаруживают участки многослойного плоского эпителия при выпадении матки, пиометре, туберкулезе и др. Чаще при этом наблюдается истинная М., осуществляющаяся с помощью резервных клеток. По данным нек-рых авторов, эпителий шейки матки при дисгормональных нарушениях может подвергаться М. в железистый призматический; эпителий желез эндометрия — в эпителий желез шейки матки и маточных труб; эпителий желез шейки матки — в эпителий желез эндометрия и маточных труб. Эпителий придатка яичка при хрон, воспалении может приобретать сходство с переходным эпителием мочевыводящих путей. М. эпителия слизистой оболочки полости рта и шейки матки в многослойный плоский ороговевающий эпителий является предопухолевым заболеванием (см.); неблагоприятным прогностическим признаком является появление очагов плоскоклеточной М. в пластах переходноклеточного рака.
Метаплазия
Классификация
Метаплазия — аномальный этап, приводящий к переформированию одного вида тканей в другой, сопровождающийся отличительными чертами функционального и морфологического плана. На данный момент, ученые выделили следующие разновидности недуга:
- Железистая — локализация поражения происходит в желудочковом, кишечном и пищеводном отделе. Подобный процесс, начинающийся в пищеводе, фиксируется в случаях предопухолевых процессах или воспалениях.
- Плоскоклеточная — протекает с заменой однослойного, призматического или цилиндрического участка эпителия на многослойный. В результате может наступить ороговение. Обычно, представленный тип болезни поражает слизистые районы организма.
Факторы, обуславливающие развитие метаплазии
Согласно медицинским данным, существует огромное число причин, провоцирующих распространение такого рода процесса. К ним относятся:
- Хронические воспалительные этапы.
- Отсутствие в достаточном количестве витамина группы А.
- Гормональные нарушения.
- Изменения, проходящие в окружающей среде.
Симптомы метаплазии
В результате поражения организма, человек начинает испытывать определенные жалобы. Все они зависят, прежде всего, от того в каком месте берут начало отклонения от нормы. В случаях проблем желудка и систем около него, пациент может ощущать:
- Тошнотные и рвотные позывы.
- Снижение уровня аппетита.
- Неприятные чувства в эпигастральном участке.
Если метаплазия образуется в цервикальной эктопии матки, что происходит только у женского пола, то больной жалуется на подобные проблемы:
- Выделения из влагалища кровяных сгустков перед месячными и после полового контакта.
- Появление отечности в нижних конечностях.
- Патологические кровоизлияния.
- Частое желание опорожнить мочевой пузырь.
В случае заражения мочевой системы, проявляются такие симптомы, как:
- Синдром гиперактивного органа.
- Едкий запах испражнений.
- Болезненные ощущения при походе в уборную.
- Регулярное чувство недостатка жидкости.
- Примеси кровеносных телец в моче.
- Жар.
Что потребуется для диагностики?
Для того, чтобы врач смог установить правильный диагноз, он проводит ряд процедур, к которым относятся:
- Беседа с пациентом, в ходе которой специалист проводит уточнение наличия симптомов и жалоб.
- Стандартные осмотр тела.
- Забор крови и мочи для тестирования.
- Микробиологическая проверка.
Читайте также: Аппликация из ткани повар
Варианты лечения
В настоящее время существует два способа избавления от метаплазии: операционное вмешательство и прием лекарственных препаратов. Используя второй путь, доктор назначает больному такие медикаменты:
- Средства антибиотического характера и стабилизирующие микрофлору кишечника.
- Вещества, направленные на профилактические меры в борьбе с онкологией.
- Компоненты для удаления рефлюксного недуга.
- Препараты для улучшения функционирования иммунитета.
Метаплазия в мезенхимальных тканях
Эпителиально-мезенхимальный переход (Epithelial-mesenchymal transition, EMT) — сложный процесс изменения эпителиальными клетками эпителиального фенотипа на мезенхимальный, происходящий в эмбриональном развиитии, заживлении ран, а также при патологических процессах — например, при фиброзе, а также при опухолевой прогрессии.
Существует также и обратный процесс — мезенхимально-эпителиальный переход (Mesenchymal-epithelial transition).
Среди механизмов EMT можно выделить несколько ключевых моментов:
- Подавление экспрессии гена Е-кадгерина (E-cadherin (CDH1)) участвующего в образовании плотных контактов между эпителиоцитами.
- Увеличение экспрессии генов ответственных за мезенхимальный фенотип эпителиоцитов, таких как виментин (Vimentin), гладко-мышечный актин, фибронектин (Fibronectin).
- Усиление клеточной подвижности вследствие активации сигнальных путей приводящих к реорганизации цитоскелета.
- Повышение экспрессии генов, кодирующих матриксные металлопротеиназы (MMP), которые участвуют в деградации внеклеточного матрикса и базальной мембраны.
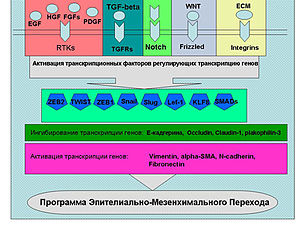
Растворимые факторы роста (на схеме), цитокины, молекулы внеклеточного матрикса активируют сигнальные пути ведущие к реализации программы ЭМП. Эти пути активируют ряд транскрипционных факторов (Snail, Twist, Slug, ZEB1, ZEB2, Lef-1 и др.), которые связываются с промоторами генов ответственных за ЭМП.
Промоторы генов, кодирующих белки плотных контактов (Tight Junction)(TJ)) — E-cadherin, occludin, claudin-1, транскрипционно ингибируются этими транскрипционными факторами, а соответственно промоторы генов компонентов цитоскелета, например Vimentin, а также генов белков внеклеточного матрикса Fibronectin, в свою очередь, наоборот активируются.
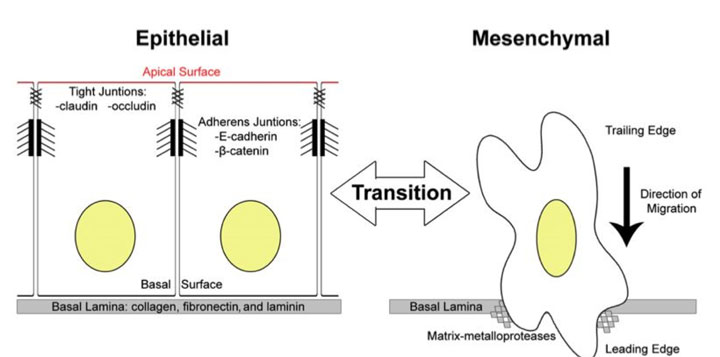
Одним из основных процессов, происходящих во время регенерации тканей является превращение эпителиальных клеток в мезенхимальные и наоборот, из мезенхимальных клеток в эпителиальные.
На рисунке выше показана разница между этими двумя основными фенотипами клеток. Важно, что нет никаких других многоклеточных тканей. Эпителиальные клетки плотно связаны друг с другом и с внеклеточным матриксом. Внеклеточный матрикс является базальной пластинкой, которая служит своего рода «колыбелью» для эпителиальных клеток. Мезенхимальные клетки расположены в 3D внеклеточной матрицы. Они биполярны, а это значит, что у них есть другое расположение цитоскелета и распределение органелл внутри них.
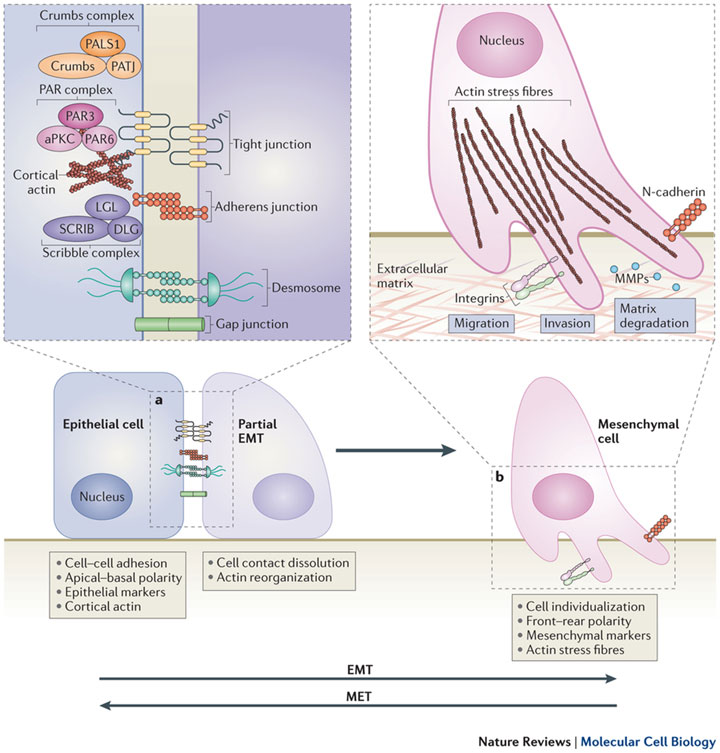
а. Первый этап эпителиально — мезенхимального перехода (epithelial-mesenchymal transition — ЕМT) – разрушение эпителиальных межклеточных контактов, плотных соединений, слипающихся между собой, десмосом и щелевых контактов , а также, нарушение клеточной полярности через нарушение частиц , разделение неисправных так называемых PAR- и Scribble (SCRIB ) -полярные комплексы. Экспрессия эпителиальных генов подавляется, одновременно с активацией экспрессии мезенхимальных генов.
б. Далее эпителиальная структура актина реорганизуется и клетки приобретают подвижность и инвазивный потенциала путем формирования ламеллиподий , филоподий и инвадопий , а также, с помощью экспрессии матричной металлопротеиназы (matrix metalloproteinases — MMPs), что, в дальнейшем, может привести к снижению внеклеточных матричных белков (extracellular matrix (ECM) proteins. Процесс мезенхимально -эпителиального перехода (МЕТ) позволяет клеткам , которые подверглись EMT вернуться в эпителий.
Утрата характерных черт эпителия (структуры ткани, клеточных взаимодействий, контроля специфическими факторами роста, приобретение подвижности и морфологии фибробластов) – это так называемое EMT, эпителиально-мезенхимальное превращение. ЕМТ свойственно нормальному эпителию в процессе развития, особенно раннего, например при гаструляции, когда эпителий приобретает подвижность и активно внедряется в подлежащие слои. ЕМТ имеет место при временных повреждениях ткани, при этом эпителиальные клетки теряют полярность, прекращают синтез кадхеринов, образуют виментин и фибронектин и одновременно с этим приобретают подвижность. Они прекращают синтез клеточных ядерных трансфакторов и образование антигенов, характерных для эпителиальных тканей. Эпителиальные клетки становятся типичными фибробластами. ЕМТ, по-видимому, лежит в основе инвазии и метастазирования: клетки эпителиальной опухоли становятся подвижными и приобретают способность расселяться по разным территориям организма. При этом очень существенно, что клетки претерпевают физиологическое, а не генетическое превращение, так как ЕМТ обратимо.
Метастазы, возникшие на основе ЕМТ, могут приобретать морфологию исходной опухоли, а эпителий в краевых районах раны может приобретать фибробластные свойства. Индукция ЕМТ имеет место при взаимодействии опухолей, экспрессирующих онкоген Ras и TGFр. Но так или иначе ЕМТ выглядит как заключительный этап прогрессии эпителиальной опухоли, когда опухоль теряет эпителиальные признаки (полярность клеток, специфические клеточные контакты, характерную морфологию и тканеспецифическую антигенную структуру) и одновременно приобретает черты фибробластов (экспрессию виментина, подвижность, независимость от территории роста).
Можно думать, что понимание этого процесса и факторов, в нем участвующих, создадут основу для рациональной терапии инвазии и метастазирования – главных свойств злокачественности. При этом непонятно, что будет дальше. Ведь прогрессия должна быть бесконечна, а EMT как бы завершает ее.
Источник: Nature Reviews Molecular Cell Biology 15,178–196, (2014)
Молекулярные основы эпителиально-мезенхимального перехода и его роль в развитии и метастазировании опухоли.

Случалось ли вам когда-нибудь желать себе обладания каким-нибудь сверхъестественным умением? Например, становиться невидимым или проходить сквозь стены? Думаю, многие не раз грезили в детстве о том, какие ловкие штуки могли бы проворачивать, будь у них какая-то эдакая способность, ставящая на порядок выше нашего мира, подчинённого обыденным устоям и правилам. Вероятно, клетки нашего организма тоже не все хотят быть такими же, как большинство, и при определённых обстоятельствах обзаводятся новыми способностями, действительно отличающими их от остальных. Но чем может обернуться такая эгоистичность для целого организма? Как вариант — зарождением опухолевого очага и стремительным распространением клеток, которые уж точно больше не серая масса, а целая гетерогенная популяция, настоящие бунтари, лишённые альтруизма и любви к своим собратьям по организму. И в сегодняшнем посте речь поведём именно о том, каким образом клетки одного типа могут принимать иную форму и какое место это событие занимает в развитии и распространении опухоли.
Читайте также: Муслин ткань какая она
Под эпителиально-мезенхимальной трансформацией (ЭМТ) понимается процесс, при котором покоящиеся эпителиальные клетки теряют свои межклеточные контакты и принимают мезенхимальную форму. Они приобретают способность к миграции через базальную мембрану, а значит, могут по кровеносному или лимфатическому руслу попасть в любые сколь угодно отдалённые от своего исходного местонахождения ткани, где путём обратной трансформации в эпителиальные клетки (мезенхимально-эпителиальный переход) формируют новые структуры. Физиологически этот процесс играет значимую роль при заживлении ран и в ходе эмбрионального развития организма. Так, например, ЭМТ лежит в основе формирования сердца, закладки большинства краниофациальных структур, а также скелетных мышц и периферической нервной системы. Да и вообще, ни один организм не разовьётся дальше стадии бластулы, не будь эпителиально-мезенхимального перехода. Однако в то же время данное явление находит своё место и в контексте патологической физиологии. ЭМТ способствует прогрессии опухолевых заболеваний за счёт того, что озлокачествленные эпителиальные клетки получают способность к проникновению в сосудистое русло, то есть, им открывается прямой путь к метастазированию. Также эпителиальные клетки, претерпев ЭМТ, могут быть задействованы в фибротических процессах: чрезмерная продукция белков внеклеточного матрикса фибробластоидными клетками может вести к функциональным нарушениям той или иной ткани.
Хотя уже в конце XIX в. эпителиальные и мезенхимальные клетки были известны как два основных типа клеток, а процесс перехода между этими двумя типами был описан в 1908 г., эпителиально-мезенхимальный переход как таковой был идентифицирован как самостоятельный процесс впервые лишь в 1982 г. Вскоре после этого было обнаружено, что если инкубировать эпителиальные клетки в среде, в которой были культивированы фибробласты, то эпителиоциты разобщаются и превращаются в мигрирующие мезенхимальные клетки. Поначалу фактор, определённый как ответственный за данную трансформацию, получил обозначение “scatter factor”, а позднее был назван фактором роста гепатоцитов (HGF), поскольку была обнаружена его роль в качестве митогена для клеток ткани печени, участвующего в регенерации печени. В экспериментах in vitro было указано на то, что HGF опосредует ЭМТ посредством активации тирозинкиназных рецепторов c-Met, кодируемых протоонкогеном. Помимо HGF и другие факторы роста осуществляют вклад в индукцию ЭМТ путём стимуляции тирозинкиназных рецепторов (как in vitro, так и in vivo), как, например, фактор роста фибробластов (FGF), инсулиноподобный фактор роста (IGF), эпителиальный фактор роста (EGF). Кроме того, большое значение придаётся трансформирующему фактору роста бета (TGF-β), который берёт на себя ключевую роль в инициации и поддержании ЭМТ как в процессе эмбрионального развития, так и при опухолевой прогрессии. Часто перечисленные факторы действуют синергетически, усиливая эпителиально-мезенхимальный переход.
Характеристика пусковых факторов и вовлечённых сигнальных путей имеет огромное значение, поскольку трансформационный процесс рассматривается непосредственно как терапевтическая мишень в контексте разработки лекарственных препаратов, направленных на борьбу с фиброзами и инвазивными опухолями. Целесообразным было бы ингибировать и пути сигнальной передачи, которые наряду с ЭМТ способствуют также ангиогенезу.
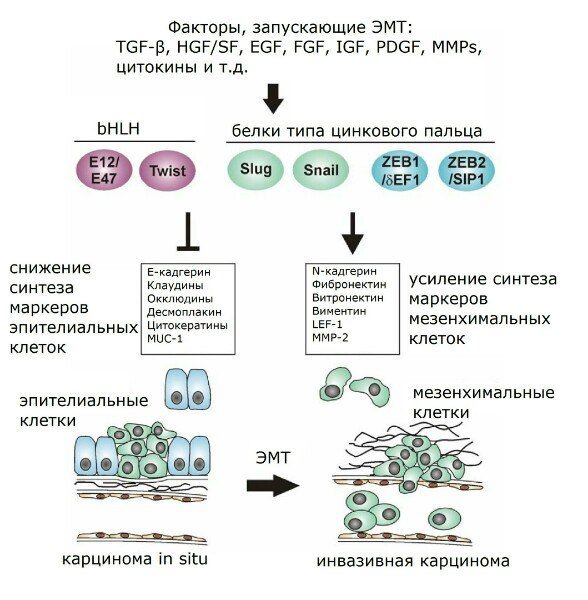
Рисунок 1 | Факторы, запускающие ЭМТ.
Итак, в ходе ЭМТ эпителиальные клетки теряют свою апикобазальную полярность и приобретают фибробластоподобный фенотип. Вследствие потери контактов с соседними клетками эпителиоциты высвобождаются из системы межклеточных связей в организованной эпителиальной ткани. После реструктуризации актинового цитоскелета клетки могут мигрировать через экстрацеллюлярный матрикс, что опосредовано тем, что на этой стадии клетки в состоянии синтезировать характерные мезенхимальные белки, дающие возможность устанавливать клеточно-матриксные связи. Также клетки вырабатывают матриксдеградирующие протеазы, разрушающие базальную мембрану и облегчающие тем самым инвазию трансформированных клеток в кровеносное и лимфатическое русло. Давайте остановимся теперь на каждом пункте чуть подробнее.
Плотные контакты (tight junctions), связывающие клетки в эпителиальных тканях, служат для поддержания целостности ткани, обеспечивают взаимодействие клеток друг с другом, а также выполняют барьерную функцию, принимая участие в регуляции проницаемости эпителия путём ограничения парацеллюлярного транспорта, и придают клеткам полярность благодаря своей организации преимущественно в апикальной области (zona occludens). Формируются плотные контакты за счёт трансмембранных белков окклюдинов и клаудинов, которые связываются с актиновым цитоскелетом через внутриклеточные белки, как например, ZO-1 (zona occludens protein 1). Адгезионные контакты (adherens junctions) представляют собой гомодимерные межклеточные контакты, характеризующиеся связью через классические кадгерины (находятся в зависимости от ионов кальция; типичные примеры — Е-кадгерин, N-кадгерин). Пронизывающие мембрану кадгерины связаны, опять же, с внутренней стороны с цитоскелетом посредством бета- и альфа-катенинов и винкулина или плакоглобина. Десмосомы, распределяющие силы напряжения по всему объёму ткани, противостоят разрывам и смещениям, то есть, важны для стабилизации ткани. Десмосомы соседних клеток связаны друг с другом за счёт трансмембранных десмосомальных кадгеринов (десмоколлины и десмоглеины), а также скреплены с кератиновыми волокнами с помощью цитоплазматических партнёров — плакоглобина и десмоплакина. Десмосомы и адгезионные контакты классификационно причисляются к заякоривающим соединениям, а плотные контакты — к запирающим.
Таким образом, при утрате того или иного соединения какой-либо клетки с окружающими содержание соответствующих контактных белков резко снижено. По этой причине эти белки годятся для трансформационного статуса эпителиальных клеток. Например, в качестве маркеров ЭМТ нередко используются многие из уже упомянутых белков — ZO-1, бета-катенин, Е-кадгерин, а также цитокератин. Снижение экспрессии или функциональной активности Е-кадгерина является крайне важной информацией о развитии опухоли, поскольку наблюдается в канцерогенезе множества злокачественных опухолей и связано с индукцией ЭМТ, а потому представляется и весьма существенным признаком инвазивного роста опухоли. Ослабление адгезионных контактов осуществляется как путём передислокации Е-кадгерина из цитоплазматической мембраны в цитоплазму, так и опосредованно через генетическую регуляцию. Белки типа цинкового пальца Snail и Slug препятствуют транскрипции Е-кадгерина, зацепляясь на специальном сайте связывания промотора гена Е-кадгерина (Е-бокс, Enhancer Box). Также Snail и Slug регулируют активацию TGF-β через Smad3. Кроме того, FGF и HGF оказывают влияние на межклеточные контакты через активирование Slug. Slug (но не Snail) принимает участие и в ЭМТ-ассоциированном заживлении ран, что совпадает с данными по различиям экспрессии гена Slug, полученными в исследованиях культуры иммортализованных нетуморогенных кератиноцитов линии НаСаТ и линии человеческих эпителиоцитов почек НК-2: если в культуре НаСаТ TGF-β индуцировал синтез только Slug, то в ренальных эпителиоцитах контроль был сосредоточен именно на белке Snail.
Читайте также: Ткани в великом новгороде ломоносова
Среди прочих факторов, оказывающих влияние на экспрессию Е-кадгерина, хотелось бы упомянуть EF1 (энхансер-связывающий фактор гамма-кристаллина), SIP1, c-Fos. EF1 и SIP1 могут управлять как транскрипцией генов белков Smad, так и, аналогично белкам Snail и Slug, напрямую взаимодействовать с промотором гена Е-кадгерина и, блокируя экспрессию Е-кадгерина, провоцировать развитие инвазивного фенотипа клетки. Эктопическая экспрессия c-Fos, например, в эпителии молочной железы, может вести к трансформации. Вообще, при кратковременной активации c-Fos потеря апикально-базальной полярности могла бы быть обратима, однако в этом случае аутокринное воздействие продуцируемого TGF-β становится инструментом поддержания ЭМТ. Так, в клетках линии НаСаТ к началу ЭМТ наблюдалась опосредованная через ERK (extracellular signal-regulated kinases) стимуляция трансформирующим фактором роста бета эктопической активности c-Fos.
Также с промотором гена Е-кадгерина способен связываться белок семейства bHLH (basic helix-loop-helix) E2A. В норме Е2А транскрипционно неактивен и представлен в виде димеров с белками ингибиторами дифференцировки (ID). В культивируемых на микроносителях эпителиальных клетках ID2 и ID3 оказываются целью воздействия TGF-β и ВМР7 (bone morphogenetic protein 7, костный морфогенетический белок): TGF-β сдерживает экспрессию ID2, вследствие чего активируется Е2А, который снижает транскрипцию гена Е-кадгерина; а ВМР7 проявляет себя антагонистически, повышая экспрессию ID2 и защищая тем самым эпителиальные клетки от трансформирующих влияний.
Важным регулятором ЭМТ как in vitro, так и in vivo проявил себя также относящийся к белкам bHLH фактор транскрипции Twist, эктопическая экспрессия которого становится причиной снижения транскрипции опять же Е-кадгерина, альфа-, бета- и гамма-катенинов. Также Twist способствует активности таких мезенхимальных маркеров, как фибронектин, виментин, альфа-гладкомышечный актин (alpha-SMA), N-кадгерин. А некоторые прочие факторы транскрипции, кроме регуляции прочности адгезионных контактов, оказывают влияние и на другие характеристики ЭМТ. Так, ID2 может препятствовать синтезу alpha-SMA; SIP1 регулирует экспрессию виментина, а в некоторых типах клеток наряду с белком Snail повышает экспрессию матриксной металлопротеиназы 2 (ММР-2); Snail может влиять на ряд существенных для ЭМТ генов (было обнаружено в клетках меланомы), среди которых гены ММР-2 и малых G-белков семейства Rho (RhoA).
Разобравшись с потерей межклеточных контактов, обратимся к тому, в чём же заключается реструктуризация цитоскелета. Для эпителиальных клеток в связи с их полярностью характерен так называемый пояс актина, волокна которого участвуют в создании адгезионных и плотных контактов, являя собой мощное дополнение периферическому актину, также задействованному в поддержании межклеточных соединений. В мезенхимальных клетках актин, напротив, представлен в виде филоподий (толстые пучки актина, цилиндроподобно вытянутые в сторону передвижения клетки), ламеллоподий (тонкие сети на переднем конце клетки, инициирующие движение клетки вперёд; придают клеточной мембране характерный внешний вид) и стрессовых волокон (пучки коротких актиновых филаментов с добавлением миозиновых филаментов; простираются параллельно продольной оси тела клетки и, сокращаясь, тоже продвигают клетку вперёд), которые поддерживают миграцию клеток. Актиновые стрессовые волокна связаны со внутренней поверхностью цитоплазматической мембраны посредством интегринов и фокальных адгезионных комплексов, имеющих в своём составе структурные белки (талин, винкулин, альфа-актинин) и некоторые протеинкиназы (например, FAK — киназа фокальных контактов). Полимеризация актина находится под контролем белков семейства Rho, куда относятся, к примеру, Rho, Rac и Cdc42, которые, приобретая активированную форму, инициируют образование как актиновых стрессовых волокон, так и адгезионных комплексов: Rac опосредует формирование ламеллоподий, Cdc42 принимает участие в образовании филоподий. Таким образом, при запуске ЭМТ белки семейства Rho представляют собой наиважнейшие целевые структуры для влияния TGF-β.
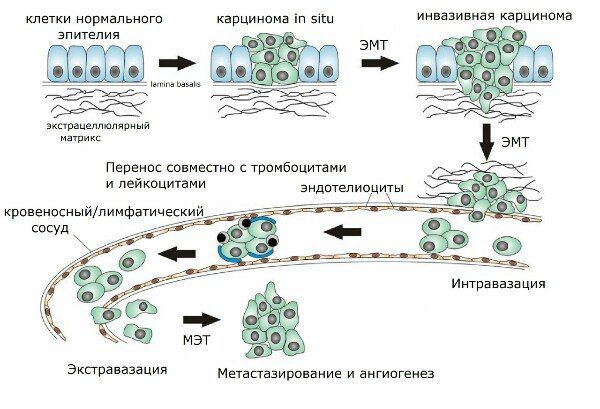
Рисунок 2 | Этапы метастазирования.
После того, как произошло переформирование внутриклеточных актиновых структур, клетка может приступать к миграции сквозь джунгли внеклеточного матрикса. Для этого ей необходимо умение образовывать новые контакты между собственными интегринами и белками матрикса. Интегрины оснащены специальными участками для связывания с матриксными белками, и благодаря RGD-последовательности интегрины распознают белки матрикса. Кроме чисто механического связывания, трансмембранные интегрины могут подвергаться контролю по типу inside-out-signalling: например, внутриклеточные киназы, связываясь с внутриклеточными участками интегринов, могут изменять их внеклеточные участки для связывания с лигандами. Но и наоборот, связывание интегринов с лигандами ведёт к внутриклеточному каскаду взаимодействий, следствием которого может быть активация FAK или белков Rho.
Ещё одна важная черта, характеризующая нормальные эпителиальные клетки — связь с базальной пластинкой посредством гемидесмосом. В ходе эпителиально-мезенхимальной трансформации клетки эпителия отделяются от базальной пластинки и начинают разрушать коллаген IV, ламинин, нидоген, из которых она построена. Для этой цели клетки вырабатывают матриксные металлопротеиназы ММР-2 и ММР-9. Структура всех ММР во многом сходна. Поначалу все ММР синтезируются как неактивные предшественники — Pro-MMPs, которые затем активируются после ферментативного отщепления продомена (эти продомены почти у всех ММР представляют собой консервативные последовательности, в которых присутствует цистеин, ответственный за связывание цинка в активном центре протеазы; поэтому только в отсутствие продомена цинк беспрепятственно связывается с активным центром и активирует протеазу). Также ММР обладают субстратспецифичными связывающими мотивами, на чём основано их разделение на подгруппы.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом