
С эмбриологической точки зрения кровяная ткань берет свое начало в первичном сосудистом ареале, мезенхимальных островках Вольфа и Пандера, от которых происходят также сердце и сосуды. В следующей главе (г) описаны фазы образования кроветворных органов, соответственно первичная мезобластичсская, затем гепатолиенальная и, на последних месяцах, окончательная костно-мозговая фаза кроветворения. Первичная мезенхимальиая клетка, содержащая информацию о кроветворной пролиферации и дифференциации восстанавливает ряды крови, как материнскими стимулами, так и стимулами, отправляемыми дифференцирующимися плодными тканями.
Привлекает внимание многосторонний потенциал первичной мезенхимальной клетки, которая порождает ретикулярную строму селезенки, узлов, печени и костей.
В послезародышевой жизни мезенхимальная клетка дифференцируется в клетки-штамм, различной направленности в костном мозге и лимфатических органах. Эта ориентация определяется в период зачаточного развития и является результатом продолжительной филогенетической эволюции. На протяжении этого развития костно-мозговая миелоидная ткань находится в зависимости от мезенхимальных, сосудисто-соединительнотканных и костных структур, составляющих частный микроклимат, способствующий дифференцированной пролиферации.
Филогенетически лимфоидная ткань развивается по плану иерархической структурной организации, причем исходно она появляется в центральных, а затем и в периферических лимфоидных органах (Берчану). Центральными органами являются вилочковая железа, у животных, и сумка Фабриция — у птиц, при том эти два органа способствуют «целенаправленности» иммунологически компетентных клеток, носящих характер лимфоцитов Т и Б. План их организации общий и создается вокруг эпителиальных образований эндотелиальной природы, берущих свое начало в пищеварительном тракте. Эти образования составляют эпителиальную ретикулярную строму, переплетающуюся с мезенхимальной стромой, как, впрочем, во всех лимфоидных органах.
Дифференциацию иммуных клеток, равно как и способность реагировать на антигенный стимул определяет их эмбриологическое происхождение, в тесной связи с эпителием, внутридермальной природы (Defendi, Good).
Функциональное различие, требуемое для дифференцированного развития по типу миелоидных или лимфоидных клеток определяется этими структурными взаимосвязями с разновидной стромой: эпителиальным эндотелием, для лимфоидной системы и костносуставными образованиями — для миелоидной. Еще не полностью выявлены взаимоотношения, налаживающиеся при создании этих двух систем. Точно установлена роль вилочковой железы в дифференциации лимфоидной системы, но не выведены размеры ее участия в функциях костного мозга. Однако из патологии известно, что опухоли вилочковой железы определяют медуллярную аплазию, но отсутствует доказательство взаимозависимости функций вилочковой железы и костного мозга при нормальном состоянии.
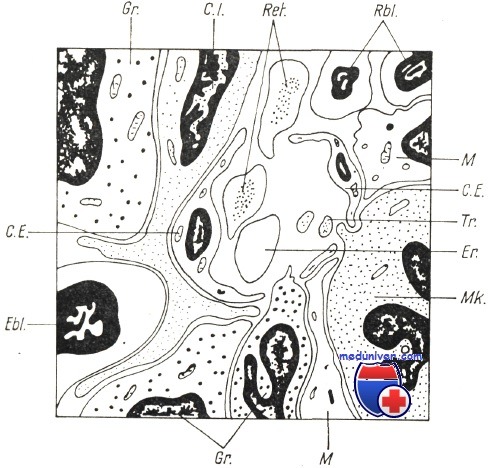
Строение костномозгового синуса.
Ebl—эритробласт; СЕ.— эндотелиальиая клетка; Gr.— гранулоцит; C.I.— интерстициальная клетка; М — макрофаг; Tr — тромбоцит; Mk — мегакариоцит; Еr— эритроцит.
На рисунке изображена функциональная взаимосвязь центральных лимфоидных органов — вилочковой железы и сумки Фабриция — и периферических лимфоидных органов — лимфатических узлов с различной топографической организацией лимфоцитов Т и Б. Лимфоциты Б, а по результатам некоторых исследований и лимфоцаты т находятся и в костном мозге, где образуют мелкие лимфоидные очаги, при этом гуморальный иммунитет находится под контролем лимфоцитов В. Поскольку морфологически клетки-штамм не отличаются от мелких лимфоцитов нам не известно являются ли костномозговые лимфоциты иммунологически компетентными клетками или клетками-штамм.
Проведенными после 1965 г. работами Yoffey и сотр. привели доказательства о том, что лимфоцит это клетка иммуного и в то же время кроветворного восстановления. Методом культур в полунепроницаемых камерах, Григориу и сотр. доказали, что периферические лимфоциты восстанавливают эритробласты. Однако последние данные, полученные в результате анализа колоний в пробирке (Metcalf u Moore) и прижизненно (Мс Colluch) выявили наличие двух различных клеток-штамм, имеющих характеристику лимфоцитов — одну из них в костном мозге, восстанавливающую миелоидные клетки, а другую в лимфоидных органах и периферической крови, восстанавливающую лимфоидные клетки. Эти аргументы выступают в пользу дуалистическоой теории о кроветворении.
Однако не ислючена мысль о способности различной дифференциации той же клетки, в зависимости от структурного, ганглиевого или медуллярного микроклимата, тем самым подтверждая неунистическую гипотезу (Loutit).
В 1979 г. Philips и сотр. впрыскивая клетки-штамм с хромосомными маркерами после облучения проследили возрождение этих клеток у других облученных животных. В связи с этим авторы заключили о наличии полипотентной клетки-штамм (КШп), из которой образуются клетка-штамм для лимфоидных линий Т и В (КШл) и клетка-штамм для миелоидной линии (КШм).
Дифференциацию по той или иной из этих линий определяют условия микроклимата и выделение специфических факторов для организации той или мной из этих линий (Boggs и сотр., Lord и сотр.). При послезародышевой жизни в остальных органах кроветворение не представляется возможным без специфического микроклимата равно как и там, где существуют угнетатели кроветворения. Однако его можно индуцировать при одновременном переносе питательной сети ретикулярной стромы (Humar и сотр.).
Читайте также: Игрушки еда из ткани
Все же функциональные связи и взаимообусловленность возможны в силу собственно структуры костного мозга, по сравнению со структурой ганглиев. В обеих структурах имеется ретикулярная строма с наличием ретикулярных и макрофаговых клеток; существуют также лимфоидные структуры, такие как паренхима ганглиев, но и лимфоидная ткань — в виде мелких фолликулов — и в костном мозге. Нам не известно выполняют ли последние и непосредственную роль в кроветворении посредством определенных общих клеток-штамм или путем стимулирующего трофического влияния на пролиферацию и дифференциацию (Берчану).
Частная патология крови выявит, что заболевания лимфоидного и миелоидного рядов различны. Глубокое поражение стромы и изменение первичной ретикулярной клетки определяет, тем не менее, нарушение обеих систем. Так, при острой недифференцированной лейкемии, рассеянной ретикулосаркоме (PC), нагрузочном ретикулезе, опухолевом гистиоцитозе поражаются обе системы с тяжелыми нарушениями и синдромами недостаточности кроветворения и иммунитета. Структурные связи в костном мозге обусловливают лимфоидные злокачественные метаплазии, смещение миелоидных клеток. При хронической лимфатической, острой лимфобластической лейкемиях или рассеянной лимфосаркоме также наблюдается смещение кроветворной ткани.
Однако существуют патологические аргументы, говорящие о дифференцированной структурной организации этих двух систем. Так, первичная или вторичная аплазия костного мозга не сопровождается аплазией лимфоидной системы; аплазия лимфоидной системы при иммунодефицитных заболеваниях в принципе не поражает кроветворную миелоидную систему. Тем не менее существуют — в настоящее время хорошо известные в иммунопатологии — комплексные иммунодефицитные заболевания, одновременно поражающие обе системы. Так, синдром ретикулярной агенезии и определенные заболевания дисгаммаглобулинемией сопровождаются одновременным нарушением иммуной лимфоидной системы и расстройствами процессов возрождения зернистых и тромбоцитных клеток (Берчану).
Функциональная взаимосвязь в норме, равно как и взаимообусловленность при патологии крови и иммуноклеточной системы говорят в пользу дифференциации миелоидной и лимфоидной систем. Однако эту дифференциацию следует рассматривать, в основном, как результат определенной степени приспособительного отбора по сравнению с условиями среды и структурного микроклимата в послезародышевой жизни. Вместе с тем организационная, генетическая и регенеративная взаимосвязи в зародышевой жизни свидетельствуют о структурном и функциональном единстве организации крови, рассматриваемой как высокодифференцированная мезенхимальная ткань.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Кроветворение во взрослом организме. Общая характеристика. Строение миелоидной и лимфоидной тканей.
К центральным органам кроветворения относятся:
1. а) Локализация — губчатое вещество плоских и губчатых костей, а также эпифизов трубчатых костей.
б) У детей (до 12-18 лет) — также диафизы трубчатых костей
(где потом красный костный мозг замещается на жёлтый костный мозг).
2. а) Консистенция — полужидкая;
поэтому из красного костного мозга приготовляют как срезы, так и мазки.
3. а) Функция: в красном костном мозгу, как говорилось выше, происходят все стадии созревания:
б) Кроме того, здесь же образуются предшественники Т-лимфоцитов, которые далее мигрируют в тимус.
Тимус (вилочковая, или зобная железа) (2)
1. Локализация — за грудиной.
2. Масса: максимальная величина — к 14-15 годам — 35-40г; затем масса органа постепенно снижается.
3. Функция: в тимусе завершается созревание Т-лимфоцитов и происходит их пролиферация, одновременно элиминируются те Т-лимфоциты, которые настроены против собственных антигенных детерминант организма.
Периферические органы кроветворения
1. Периферические органы кроветворения составляют т.н. периферическую лимфоидную систему, которая включает: лимфоидную систему слизистых оболочек, многочисленные лимфатические узлы (4), располагающиеся по ходу лимфатических сосудов, и селезёнку.
2. Общая масса лимфоидной ткани во всех этих образованиях сравнима с массой печени или головного мозга.
3. Очень многочисленны компоненты лимфоидной системы слизистых оболочек:
а) глоточное лимфоидное кольцо (или кольцо Пирогова) –
одиночные (солитарные) лимфатические фолликулы, а также их скопления (пейеровы бляшки)
в) в стенке червеобразного отростка — лимфатические узелки (3.Г),
г) в стенке воздухоносных путей — лимфатические узелки (бронхоассоциированная лимфоидная ткань — БАЛТ).
а) В периферической лимфоидной ткани, как уже отмечалось, оседают В- и Т-лимфоциты из центральных органов кроветворения, образуя лимфоидные узелки.
б) Именно здесь происходит встреча лимфоцитов (В- и Т-клеток) с антигенами — чужеродными молекулами (которые могут находиться либо в растворённом состоянии, либо на поверхности клеток).
в) Это вызывает соответствующие иммунные реакции, которые обычно включают и интенсивную пролиферацию антигенстимулированных клеток.
I. Два типа кроветворения — миело- и лимфопоэз
1. Из приведённого описания видно:
образование лимфоцитов (отвечающих за иммунный ответ) стоит несколько особняком от образования других клеток крови(хотя и то, и другое начинается в одном органе — красном костном мозгу).
Читайте также: Авто ткань в курске
2. Соответственно, выделяют два вида кроветворения:
а) миелопоэз — образование всех форменных элементов крови, кроме лимфоцитов, т.е. эритроцитов, гранулоцитов, моноцитов и тромбоцитов;
б) лимфопоэз — образование лимфоцитов (Т- и В-клеток).
II. Два типа кроветворной ткани — миелоидная и лимфоидная
1. а) Ткань, в которой происходит миелопоэз, называется миелоидной.
б) Это — красный костный мозг.
в) Но, как мы уже знаем, в миелоидной ткани, кроме миелопоэза, совершаются и важные события лимфопоэза:
созревание В-лимфоцитов и начальные стадии созревания Т-лимфоцитов.
2. а) Ткань, в которой происходит дозревание и функционирование лимфоцитов, называется лимфоидной.
III. Два компонента кроветворной ткани — стромальный и гемальный
И в миелоидной ткани костного мозга, и в лимфоидной ткани соответствующих органов содержатся два основных компонента.
1. Первый — стромальный компонент. Он может быть представлен:
ретикулярной тканью- в красном костном мозгу, лимфоузлах и селезёнке,
рыхлой соединительной тканью — в лимфатических фолликулах слизистых оболочек,
эпителиальной тканью — в тимусе.
2. а) Второй компонент — гемальный:
гемопоэтические (кроветворные) клетки на разных стадиях созревания.
б) Они находятся в тесной связи с элементами стромального компонента, образующими микроокружение.
IV. Происхождение кроветворной ткани
1. О мезенхимном происхождении кроветворных клеток нам уже известно (п. 20.1.1.1).
2. а) Клетки стромального компонента (будучи разновидностью соединительной ткани) тоже имеют такое же происхождение.
б) Исключение, как уже отмечалось, составляет тимус:
здесь строма долек представлена эпителиальной тканью.
Образование эритроцитов.
Формирование эритроцитов Формирование эритроцитов (эритропоэз) происходит в красном костном мозге тазовых костей, черепа, рёбер и позвоночника, а у детей — ещё и в костном мозге в окончаниях длинных костей рук и ног. В процессе созревания они теряют ядро и становятся безъядерными. Продолжительность жизни эритроцита — 3—4 месяца, разрушение (гемолиз) происходит в печени и селезёнке.
Внутри эритроцитов находятся молекулы ярко-красного дыхательного пигмента — гемоглобина, который делает кровь красной.
Гемоглобин состоит из двух частей: белковой — глобина — и железосодержащей — гема.
Гемоглобин способен легко присоединять кислород. Соединение гемоглобина с кислородом имеет ярко-красный цвет. Кровь, насыщенную кислородом, называют артериальной.
Соединение гемоглобина с кислородом нестойкое. При его распаде вновь образуются гемоглобин и свободный кислород, который поступает в клетки тканей. Кровь, обеднённую кислородом, называют венозной.
Функция эритроцитов — дыхательная (доставка к тканям кислорода и удаление углекислого газа).
Миелоидная ткань где находится
Эмбриональный источник развития кроветворных тканей — мезенхима. В эмбриогенезе можно различать 3 периода гемопоэза: 1) внезародышевый, или мезобластический (1-2 мес), 2) гепато-тимо-лиенальный (2-5 мес), 3) медулло-тимо-лимфоидный (5-10 мес).
Постэмбриональным гемопоэзом, или физиологической регенерацией крови, называют кроветворение во взрослом организме.
Кроветворение в желточном мешке. В конце 2-й — начале 3-й недели эмбриогенеза в мезенхиме стенки желточного мешка образуются кровяные островки, в составе которых клетки дифференцируются на плоские эндотелиальные и округлые клетки. Последние преобразуются в стволовые кроветворные клетки. При внезародышевом кроветворении из стволовых клеток формируются первичные эритробласты — мегалобласты. Они делятся внутри сосудистого русла (интраваскулярно). Небольшая часть мегалобластов превращается в безъядерные первичные эритроциты — мегалоциты. Образуется также незначительное количество вторичных эритроцитов меньшей величины, чем мегалоциты. Экстраваскулярно дифференцируется часть первичных лейкоцитов (гранулоцитов — нейтрофилов и эозинофилов). Из желточного мешка стволовые кроветворные клетки по развивающимся сосудам расселяются по органам зародыша.
Кроветворение в печени. На 2-м месяце эмбриогенеза печень становится центром кроветворения. Источником гемопоэза здесь служат стволовые кроветворные клетки. Кроветворение в печени происходит экстраваскулярно. Из стволовых кроветворных клеток образуются эритроциты, зернистые гранулоциты (нейтрофилы и эозинофилы) и мегакариоциты. Зернистые лейкоциты развиваются здесь укороченным путем и не имеют четкой специфической зернистости. К концу эмбриогенеза человека кроветворение в печени постепенно прекращается.
Кроветворение в селезенке. На 4—5-м месяцах эмбриогенеза человека селезенка становится универсальным органом гемопоэза, в котором экстраваскулярно образуются все клетки крови. Позднее процессы эритро- и гранулоцитопоэза в селезенке угасают, но усиливается образование незернистых лейкоцитов.
Кроветворение в красном костном мозге и тимусе. Постепенно центральным органом кроветворения становится красный костный мозг. Строму его вначале образует мезенхима, которая позднее преобразуется в ретикулярную ткань. Ретикулярная ткань, в трехмерной сети которой происходит развитие эритроцитов, гранулоцитов, моноцитов и мегакариоцитов, называют миелоидной тканью. Миелоидная ткань — специализированная гемопоэтическая ткань красного костного мозга. Она обеспечивает развитие стволовых клеток и всех форменных элементов крови. Наряду с миелоидной к кроветворным тканям относится лимфоидная ткань, которая развивается в лимфатических узлах, селезенке и других лимфоидных органах, составляющих лимфоидную систему. Здесь в сети ретикулярной ткани происходит образование лимфоцитов, плазматических клеток, удаление клеток и продуктов их распада.
Читайте также: Пластмассовая линейка натертая шерстяной тканью получила положительный заряд
К центральным органам кроветворения относится тимус, в котором на 2-м месяце эмбриогенеза начинают дифференцироваться лимфоциты тимуса. В дальнейшем они расселяются по периферическим органам лимфоидной системы.
Кроветворение в лимфатических узлах начинается с 4-го месяца эмбриогенеза после миграции стволовых кроветворных клеток.
В соответствии с унитарной теорией кроветворения А.А. Максимова, существует единый источник развития для всех клеток крови. Исходной клеткой для всех ростков кроветворения является стволовая кроветворная клетка, сходная по своему строению с малым лимфоцитом. А.А. Максимов (1911) писал, что индифферентные блуждающие клетки, или лимфоциты в широком смысле, одарены очень большой потенцией развития: «Это индифферентная мезенхимная блуждающая клетка, лимфоцит, является общей родоначальницей всех элементов крови. Попадая в благоприятные условия, она проявляет свою потенцию развития, причем в зависимости от условий, направление развития и продукты его получаются очень разнообразными». Унитарная теория кроветворения была развита в трудах А.А. Заварзина, Н.Г. Хлопина, А.Н. Крюкова, М.И. Аринкина и др. Метод селезеночных колоний, разработанный канадскими учеными Тиллом и МакКуллохом (1961), прозволил идентифицировать вид клеток, являющийся источником развития клеток эритроидного, гранулоцитарного и мегакариоцитарного рядов. Эту клетку, которая гистологически сходна с малым темным лимфоцитом, авторы назвали колониеобразующей единицей (КОЕ).
В развитии клеток крови условно выделяются классы клеток. По мере перехода клеток из класса в класс, в каждом из них все более отчетливо обнаруживаются гемопоэтические клеточные диффероны, которые характеризуются определенными гистологическими признаками. Однако клетки первых трех классов по своему строению идентичны. Только методы иммуноцитохимии позволяют различать клетки по наборам клеточных рецепторов, что является показателем дивергентной дифференцировки стволовой клетки.
В общем виде развитие клеток крови происходит в следующей последовательности.
1-й класс — плюрипотентные клетки — это стволовые кроветворные клетки (СКК). Стволовая клетка является общим самоподдерживающимся предшественником всех клеток крови, включая все виды иммунокомпетентных клеток. Полагают, что каждая из стволовых клеток способна проделать по меньшей мере 100 митозов, т. е. потомками одной стволовой клетки можно было бы обеспечить всю кроветворную систему. Однако стволовые клетки после цикла пролиферации в эмбриогенезе переходят в состояние покоя. Она лишена каких-либо специфических признаков строения и локализуется в миелоидной ткани среди популяции лимфоците- или моноцитоподобных элементов. Она может с током крови мигрировать по тканям организма. Объективным методом обнаружения и количественного учета стволовых клеток является метод селезеночных колоний. Стволовые клетки составляют около 0,1% популяции кроветворных элементов.
2-й класс. Стволовые клетки под влиянием ряда факторов (тромбопоэтический, ИЛ-7 и др.) дивергентно дифференцируются в двух направлениях: полустволовые, или мультипотентные, клетки — предшественники миелопоэза (КОЕ-ГЭММ) и мультипотентные клетки — предшественники лимфопоэза (КОЕ-Л). В составе колоний эти клетки имеют ограниченные возможности к самоподдержанию (около 3-4 недель), однако этого достаточно для поддержания физиологической регенерации крови.
3-й класс. Из мультипотентных клеток — предшественников миелопоэза (КОЕ-ГЭММ) в результате дивергентной дифференцировки, происходящей под влиянием ряда факторов микроокружения, возникают следующие клеточные линии: а) родоначальные (прогениторные) клетки, или клетки-предшественники, эритропоэза (БОЭ-Э, от англ. burst — взрыв) и развивающиеся из них КОЕ-Э; б) общие родоначальные клетки гранулоцито- и моноцитопоэза (КОЕ-ГМо). Последние в процессе дальнейшей дивергентной дифференцировки под влиянием факторов микроокружения формируют родоначальные клетки для нейтрофильных (гранулоцит-стимулирующий фактор), эозинофильных (ИЛ-5) и базофильных (ИЛ-3) гранулоцитов (КОЕ-Гн, КОЕ-Эо, КОЕ-Б) и моноцитов (КОЕ-Мо, фактор — моноцит-колониестимулирующий).
Мультипотентные клетки лимфопоэза (КОЕ-Л) под влиянием дифференцировочных факторов микроокружения (ИЛ-7, ИЛ-6) развиваются в родоначальные клетки Т- и В-лимфоцитов.
Мультипотентные клетки КОЕ-ГЭММ (при участии тромбопоэтина и ИЛ-11) являются источником развития родоначальной клетки для мегакариоцитов (КОЕ-Мег).
Таким образом, важнейшее свойство, которое приобретают в миелопоэзе и лимфопоэзе кровеобразующие клетки — это формирование рецепторно-трансдукторной системы, реагирующей на конкретные факторы дифференцировки (эритропоэтин, тромбопоэтин, колониестимулирующие факторы, интерлейкины — ИЛ и др.), вырабатываемые кроветворным микроокружением и клетками других органов. Все это приводит к тому, что в клетках появляются гистологические маркеры, на основе которых можно с большой вероятностью отнести ту или иную клетку к конкретному гемопоэтическому ряду (дифферону).
IV-й класс клеток — гистологически распознаваемые клетки кроветворной ткани — это пролиферирующие клетки («бласты»). Они способны к пролиферации и дифференцировке.
V-й класс — созревающие клетки («про-циты») и VI класс — зрелые клетки периферической крови.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом