
Минерализация органического матрикса остеобластами осуществляется двумя основными механизмами:
(1) путем отложения кристаллов гидроксиапатита из перенасыщенной внеклеточной жидкости вдоль фибрилл коллагена. Секретируемые остеобластами неколлагеновые белки контролируют ход минерализации.
(2) посредством секреции особых матричных пузырьков — мембранных структур, которые выделяются в матрикс остеобластами. В них содержатся высокие концентрации фосфата кальция и щелочной фосфатазы и др. Разрушаясь, пузырьки служат ядрами, вокруг которых растут кристаллы гидоксиапатита. В дальнейшем очаги минерализации увеличиваются в размерах и сливаются друг с другом, превращая новообразованный остеоид в зрелый костный матрикс.
В результате минерализации 90-95% солей кальция включаются в состав коллагеновых волокон и лишь 5-10% находятся в остальной части матрикса.
В норме минерализация осуществляется вскоре после образования остеоида, занимая у человека примерно 15 сут.
Нарушение процессов минерализации кости происходит при снижении в крови уровня кальция (вследствие недостаточного поступления с пищей, нарушения всасывания) или фосфата (обычно при усиленном выделении с мочой).
Кальций в кристаллах гидроксиапатита может замещаться другими элементами; наиболее опасно его замещение радиоактивными стронцием ( 90 Sr) плутонием ( 259 Ри) или другими продуктами расщепления урана. Эти элементы могут попадать в костную ткань из внешней среды при ее радиоактивном заражении. Включаясь в состав костной ткани, и длительно в ней находясь, они вызывают сильное внутреннее облучение организма, повреждая, в первую очередь, костный мозг. В результате развивается хроническая лучевая болезнь, саркома кости.
Регуляция деятельности остеобластов осуществляется гормонами и другими биологически активными веществами благодаря наличию на их плазмолемме специфических рецепторов паратгормона, витамина D, глюкокортикоидов, половых гормонов (андрогенов, эстрогенов), кальцитонина, тиреоидных гормонов, факторов роста, инсулина, простагландинов. Они реагируют также на факторы, продуцируемые остеокластами, и секретируют вещества, обусловливающие их собственную активацию (аутокринные регуляторы).
Остеоциты – основной тип клеток зрелой костной ткани. Остеоциты — зрелые и уже не делящиеся клетки, расположенные в костных полостях, или лакунах (рис. 6.6.). В клетке присутствуют цистерны гранулярной эндоплазматической сети, свободные рибосомы, комплекс Гольджи, округлые митохондрии и лизосомы. Тонкие отростки остеоцитов расположены в канальцах, отходящих в разные стороны от костных полостей. Отростки соседних остеоцитов, соприкасающиеся боковыми поверхностями, формируют щелевые контакты. Совокупность канальцев и лакун образует – лакунарно-канальцевую систему (6. 6).
Она заполнена тканевой жидкостью, через которую осуществляется обмен между остеоцитами и кровью. Разделяющий плазму и лакунарно-канальцевую жидкость, барьер называют костной мембраной. Барьер формируют остеобласты и остеоциты. Концентрация Са 2+ и РО4 -з в лакунарно-канальцевой жидкости превышает критический уровень для спонтанного осаждения солей Са 2+ , что указывает на присутствие и важную роль различных ингибиторов осаждения контролирующих процесс минерализации.
Функция остеоцитов состоит в поддержании нормального состояния костного матрикса (и баланса Са и Р в организме). При этом они не только вырабатывают его компоненты, но, по-видимому, обладают способностью к ограниченному растворению матрикса, что приводит к увеличению объема лакун (остеоцитарный остеолиз). Это явление у здоровых людей отмечается в 3-4% лакун; оно усиливается в несколько раз при повышенных уровнях паратгормона или недостатке витамина D. Остеоциты воспринимают механические напряжения, возникающие внутри костной ткани; они, очевидно, чувствительны и к электрическим потенциалам, образующимся в матриксе при воздействии деформирующих сил. Реагируя на эти и другие сигналы, остеоциты запускают локальный процесс перестройки костной ткани, ограниченный мелким участком скелета.
Остеокласты – многоядерные содержат (20-50 ядер) гигантские (20-100 мкм) клетки (точнее говоря, симпластические структуры, образующиеся вследствие слияния моноцитов), обладающие подвижностью и осуществляющие разрушение, или резорбцию (от лат. resorptio – рассасывание) костной ткани. Они располагаются в образованных ими углублениях на поверхности костной ткани (резорбционных лакунах, или лакунах Хаушипа) поодиночке или небольшими группами (рис. 6.7.), способны проделывать в костной ткани глубокие ходы (тоннели).
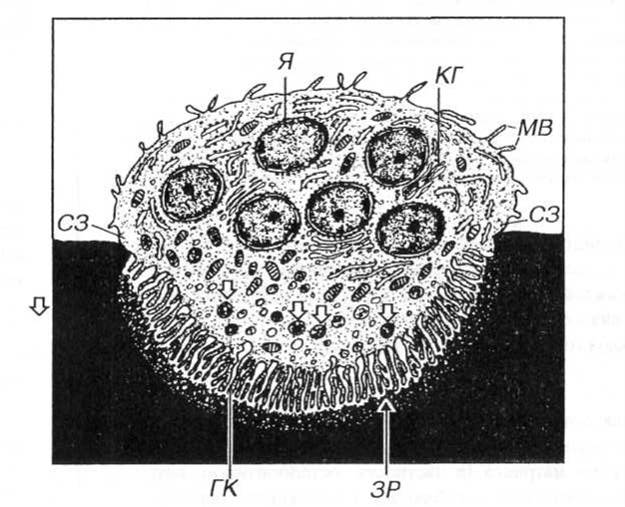
О стеокласты – резко поляризованные клетки. В активном остеокласте участок его цитоплазмы, прилежащий к кости и не содержащий ядер и большинства органелл, образует многочисленные складки клеточной мембраны (гофрированный край). В отличие от исчерченной каемки, состоящей из микроворсинок, выпячивания цитоплазмы остеокласта в области гофрированного края — вариабельные структуры, постоянно вытягивающиеся и сокращающиеся. По обеим сторонам гофрированного края имеются гладкие краевые светлые зоны — участки плотного прикрепления его цитоплазмы к кости.
Рис. 6.7. Ультраструктурная организация остеокласта. Схема. Цитоплазма остеокласта образует многочисленные складки клеточной мембраны — гофрированный край (ГК) — участок обеспечивающий резорбцию кости. Резорбция включает деминерализацию матрикса в зоне резорбции (ЗР) и переваривание его органических компонентов в лизосомах (стрелки). Плотное прикрепление цитоплазмы остеокласта к кости осуществляется в области краевых светлых зон (СЗ). MB — микроворсинки, КГ -комплекс Гольджи. Я — ядро.

Р ис. 6.8. Механизм резорбции костной ткани остеокластом. Остеокласт прикрепляется к поверхности кости в участке резорбции; особо плотное прикрепление образуется в области краевых светлых зон (СЗ). Закисление содержимого резорбционной лакуны (РЛ), обусловливающее растворение минерального компонента матрикса, осуществляется путем экзоцитоза пузырьков с кислым содержимым (ПКС), сливающихся с плазмолеммой остеокласта в области гофрированного фая (ГК),’ а также благодаря действию протонных насосов (АТФазы мембраны ГК), накачивающих ионы Н в РЛ. Источником протонов служит реакция между СО2, и Н2О, катализируемая ферментом карбоангидразой (КА). Органические компоненты матрикса разрушаются лизосомальными ферментами, выделяемыми в лакуну. Продукты резорбции костной ткани удаляются из лакуны путем их утечки (У) в области СЗ (механизм «разгерметизации» лакуны) или везикулярным транспортом (ВТ) через цитоплазму клетки.
Читайте также: Абажуры из ткани классика
Резорбции костной ткани остеокластами протекает циклически (рис. 6.8.).
1) прикрепление остеокластов к резорбируемой поверхности кости обеспечивается рядом адгезивных взаимодействий, опосредованных интегринами и белками матрикса (в частности, остеопонтином, витронектином). При этом в остеокласте наблюдается выраженная перестройка элементов цитоскелета. Особое плотное прикрепление к костной ткани остеокласты образуют в области краевых светлых зон, тем самым «герметизируя» зону резорбции, и препятствуя в дальнейшем утечке из нее протонов;
2) закисление содержимого лакун осуществляется двумя механизмами: (а) путем выделения кислого содержимого вакуолей в лакуну; (б) благодаря действию протонных насосов (АТФазы мембраны гофрированного края), накачивающих ионы Н + в лакуну;
3) резорбцию минерального компонента матрикса, которая осуществляется вследствие воздействия на него кислого содержимого лакуны;
4) растворение органических компонентов матрикса вследствие действия лизосомальных ферментов остеокластов, секретированных ими в лакуну и активирующихся при низких значениях рН. Высказывается мнение о том, что остеокласты осуществляют лишь деминерализацию матрикса, а разрушение органических компонентов обеспечивается макрофагами;
5) удаление продуктов разрушения костной ткани осуществляется двумя механизмами: (а) их утечкой из лакуны после отделения плазмолеммы от поверхности кости (механизм «разгерметизации» лакуны), (б) поглощением продуктов остеокластами, и их везикулярным транспортом через цитоплазму клетки с последующим выделением в области ее апикального полюса.
Так как резорбция кости сопровождается освобождением связанного с ее матриксом кальция, эти клетки играют важнейшую роль в поддержании кальциевого гомеостаза.
Регуляция активности остеокластов осуществляется гормоном околощитовидных желез (па-ратгормон), 1,25 гидроксивитамин D3 (активируют остеокласты и увеличивают их число, стимулируя слияние мононуклеарных предшественников). Гормон щитовидной железы кальцитонин и женские половые гормоны (эстрогены) угнетают активность остеокластов. Кальцитонин связывается со специфическими рецепторами на поверхности остеокластов, а параттормон, рецепторы которого на остеокластах отсутствуют, оказывает на них непрямое действие, повидимому, опосредованное остеобластами.
Метаболизм костной ткани и остеопороз
Рассмотрены подходы к выбору средств для профилактики и лечения потерь костной ткани, восстановления ее структуры и качества. Применяемый препарат должен способствовать синтезу коллагена, формированию костного матрикса, его минерализации и, соответственно
Approaches to selecting the methods of prevention and treatment of bone tissue losses, restoration of its structure and quality, were considered. The used preparation must contribute to collagen synthesis, formation of bone matrix, its mineralization, and, respectively, increase of the bone density and strength.
Остеопороз (ОП) — прогрессирующее системное заболевание скелета, характеризующееся снижением костной массы и нарушением микроархитектоники (качества) костной ткани, что приводит к хрупкости костей и повышению риска переломов. ОП — самое распространенное заболевание костной ткани: остеопоротические переломы отмечается у половины всех женщин, находящихся в периоде постменопаузы, а также у мужчин старших возрастных групп [1]. Очевидно, что рано начатые активные профилактические мероприятия у значительной части населения могут существенно повлиять на распространенность, прогрессирование и исходы заболевания, а также снизить риск переломов. В связи с этим изучение различных лекарственных препаратов и методов, применяемых для профилактики ОП, приобретает особый смысл.
Кость — специализированная разновидность соединительной ткани, состоящая из клеток и межклеточного вещества. В течение всей жизни основные функции костной ткани, такие как жесткость и гибкость, снижаются, поскольку с возрастом наблюдаются повреждение матрикса и потеря минералов. В противовес указанным проявлениям, в кости осуществляется ремоделирование — процесс, направленный на самостоятельное обновление и сохранение скелета как структурного и функционального органа.
Основными клетками костной ткани, функциями которой регулируется гомеостаз кости, являются остеобласты, остеокласты и остеоциты. Основной функцией остеобластов является создание органического межклеточного матрикса кости, остеоида. Остеобласты синтезируют и выделяют в окружающую среду фибриллы коллагена, протеогликаны и гликозаминогликаны. Наряду с этим остеобласты активно синтезируют и выделяют во внеклеточное пространство значительное количество глицерофосфолипидов, способствующих связыванию Ca 2+ и участвующих в процессах минерализации. Клетки сообщаются между собой через десмосомы, которые позволяют проходить Ca 2+ и цАМФ. Они также обеспечивают непрерывный рост кристаллов гидроксиапатитов и выступают в качестве посредников при связывании минеральных кристаллов с белковой матрицей.
В ходе формирования кости некоторые остеобласты оказываются замурованными в толщу матрикса и становятся остеоцитами. Остеоциты контактируют друг с другом через отростки, являются основными компонентами в сформировавшейся костной ткани. Основная функция остеоцитов — поддержание нормального состояния костного матрикса и баланса кальция и фосфора в организме.
Остеокласты — клетки, выполняющие функцию разрушения кости; развиваются из стволовой кроветворной клетки и являются специализированными макрофагами. В процессе ремоделирования кости резорбтивный стимул запускает процесс привлечения остеокластов к участку кости. Прикрепившись к кости, остеокласты продуцируют множество протеолитических ферментов и формируют полость в кальцинированном матриксе. Таким образом, они осуществляют непрерывный процесс резорбции и обновления костной ткани, обеспечивая необходимый рост и развитие скелета, структуру, прочность и упругость.
Важнейшим компонентом костной ткани является межклеточное вещество — уникальный комплекс органических и неорганических компонентов, заполняющих пространство между клетками. Минерализованный матрикс костной ткани поддерживает структуру скелета и под координирующим влиянием остеобластов и остеокластов обеспечивает резервуар как ионов, так и факторов роста, которые высвобождаются в процессе метаболизма.
Органический межклеточный матрикс костной ткани представлен семейством коллагеновых белков. Состав кости необычен тем, что фактически в ней представлен только коллаген I типа (90%), хотя наряду с коллагеном I типа в кости все же присутствуют следы других типов коллагена, таких как V, XI, XII. Скорее всего, что эти типы коллагена принадлежат другим тканям, которые и находятся в костной ткани, но не входят в состав костного матрикса. Например, коллаген V типа обычно обнаруживается в сосудах, которые пронизывают кость. Коллаген XI типа находится в хрящевой ткани и может соответствовать остаткам кальцифицированного хряща. Коллагеновые фибриллы в кости строго ориентированы в соответствии с распределенной функциональной нагрузкой на кость, что обеспечивает упругость и эластичность кости. Веретенообразные и пластинчатые кристаллы гидроксиапатита находятся на коллагеновых волокнах, в их пределах и в окружающем пространстве. Как правило, они ориентированы в том же направлении, что и коллагеновые волокна.
Читайте также: Ткани анатомия таблица функции
Неколлагеновая часть матрикса (10%) представлена основным веществом (витамин К-зависимыми глютамилпротеинами (остеокальцином), матричными протеинами, остеопонтином, остеонектином, фибронектином, фосфопротеидами, сиалопротеидами, а также протеогликанами).
Минеральные вещества, которыми пропитан органический матрикс, представлены главным образом кристаллами гидроксиапатита Ca10(PO4)6(OH)2. Кроме того, в кости обнаружены ионы Mg 2+ , Na + , K + , SO4 2- , HCO 3- , гидроксильные и другие ионы, которые могут принимать участие в образовании кристаллов.
Важно подчеркнуть, что ОП является результатом уменьшения органического матрикса кости, а вовсе не плохой кальцификацией костной ткани. При ОП существенно снижается скорость образования остеоида, необходимого для формирования кости. Поэтому при планировании профилактических мероприятий чрезвычайно важно учитывать потенциальную возможность препаратов, наряду с адекватной минерализацией, оказывать влияние на синтез органического матрикса.
Разумеется, качественная структура и прочность кости, ее эффективное функционирование и своевременное самообновление возможны лишь при адекватной обеспеченности макро- и микроэлементами, которые, подобно кальцию и витамину D, принимают непосредственное участие в биохимических процессах костной ткани [2–5]. Магний, медь, цинк, марганец, бор, являясь кофакторами ферментов, регулируют синтез костного матрикса, его минерализацию, а также равномерный рост, гибкость и прочность костной ткани. Известно, что дефицит этих веществ замедляет формирование костной массы в детстве и подростковом возрасте, способствует ее ускоренной потере в пожилом возрасте. Соответственно, дефицит любого из известных минеральных веществ в организме препятствует успешной терапии и профилактике нарушений структуры кости [6, 7].
Одним из основных минералов, играющих важную роль в формировании и поддержании структуры костной ткани, является кальций. Поскольку кальций не производится в организме, то для поддержания оптимальной концентрации он должен регулярно поступать извне. Причем желательно, чтобы его поступление в организм обеспечивалось за счет натуральных молочных продуктов, молока и его производных (кефира, простокваши, ряженки, йогурта, творога, сыра). Вместе с тем биодоступность кальция из пищи составляет порядка 30%, причем с высокой индивидуальной вариабельностью. Более того, у лиц пожилого возраста нередко имеет место непереносимость молочных продуктов, связанная со снижением концентрации лактазы в желудочном соке, что приводит к низкому потреблению кальция.
Согласно эпидемиологическим исследованиям, среди женщин в возрасте старше 45 лет, проживающих в мегаполисах, непереносимость молока встречается с частотой 25,0–34,0%. При этом достаточное потребление кальция с продуктами питания имеет место менее чем у 5% женщин [8]. Фактически содержание кальция в пищевом рационе постменопаузальных женщин не соответствует рекомендованным нормам. Очевидно, что обеспечение должного уровня потребления кальция возможно лишь при условии дополнительного регулярного назначения медикаментозных препаратов.
Витамин D — основной регулятор активной абсорбции кальция в организме. Витамин D относят к группе жирорастворимых витаминов. Хотя в отличие от всех других витаминов он биологически не активен. В активную, гормональную, форму он превращается за счет двухступенчатой метаболизации в организме и оказывает многообразные биологические эффекты за счет взаимодействия со специфическими рецепторами, локализованными в ядрах клеток тканей и органов. Другое дело — активный метаболит витамина D. Он действует как истинный гормон, хотя в научной литературе его традиционно называют витамином D [9, 10].
Природная форма витамина D — витамин D2 (эргокальциферол) поступает в организм человека в относительно небольших количествах — не более 20–30% от потребности. В основном из злаковых растений, рыбьего жира, сливочного масла, маргарина, молока, яичного желтка и др. В организме витамин D2 метаболизируется с образованием производных, обладающих сходным с метаболитами витамина D3 действием.
Еще одна природная форма витамина D — витамин D3, или холекальциферол, является ближайшим аналогом витамина D2, но его синтез мало зависит от поступления извне. Холекальциферол образуется в организме позвоночных животных, в том числе амфибий, рептилий, птиц и млекопитающих, в связи с чем играет значительно бóльшую роль в процессах жизнедеятельности человека, чем поступающий в небольших количествах с пищей витамин D2. В организме витамин D3 образуется из находящегося в дермальном слое кожи предшественника (7-дегидрохолестерина) под влиянием коротковолнового ультрафиолетового облучения спектра В (УФ–В/солнечного света, длина волны 290–315 нм) при температуре тела в результате фотохимической реакции раскрытия В-кольца стероидного ядра и термоизомеризации, характерной для секостероидов [9, 10].
В последующем поступивший с пищей и/или образовавшийся в организме в процессе эндогенного синтеза витамин D подвергается реакции 25-гидроксилирования в печени. Важно, что гидроксилирование витамина D3 в печени представляет собой полностью субстратзависимый процесс, который протекает весьма быстро и ведет к повышению уровня 25(ОН)D в сыворотке крови. Уровень этого вещества отражает как образование витамина D в коже, так и его поступление с пищей, в связи с чем может использоваться как маркер статуса витамина D [9, 10].
Читайте также: Какую ткань выбрать для штор в гостиную лучше
Вторая реакция гидроксилирования 25(ОН)D, с образованием наиболее важной, качественно и количественно значимой активной гормональной формы — 1a,25-дигидроксивитамина D3 (1α,25(ОН)2D3), называемой также D-гормоном, кальцитриолом, протекает уже в основном в почках, в клетках проксимальных отделов канальцев коры почек при участии фермента 1α-гидроксилазы (CYP27В1). Этот процесс строго регулируется рядом эндогенных и экзогенных факторов. Во-первых, регуляция синтеза 1a,25(ОН)2D3 в почках является непосредственной функцией паратиреоидного гормона (ПТГ), на концентрацию которого в крови, в свою очередь, по механизму обратной связи оказывают влияние как уровень самого активного метаболита витамина D3, так и концентрация кальция и фосфора в плазме крови. Во-вторых, активация синтеза 1a-гидроксилазы и реакции 1a-гидроксилирования зависит от половых гормонов (эстрогенов и андрогенов), кальцитонина, пролактина, гормона роста (через ИПФР-1) и др. В-третьих, ингибирующее влияние на активность 1a-гидроксилазы оказывают глюкокортикостероидные гормоны, 1α,25(ОН)2D3 и ряд его синтетических аналогов. Фактор роста из фибробластов (FGF23), секретируемый в клетках кости, вызывает образование натрий-фосфат-котранспортера, который действует в клетках почек и тонкого кишечника, оказывает тормозящее влияние на синтез 1,25-дигидроксивитамина D3. На метаболизм витамина D оказывают влияние и некоторые лекарственные средства, например, противоэпилептические препараты.
Основными реакциями, в которых участвует D-гормон, являются абсорбция кальция в желудочно-кишечном тракте и его реабсорбция в почках. D-гормон усиливает кишечную абсорбцию кальция в тонком кишечнике за счет взаимодействия со специфическими РВD. Об эффективности данного механизма свидетельствует тот факт, что без участия витамина D лишь 10–15% пищевого кальция и 60% фосфора абсорбируются в кишечнике. Взаимодействие между 1a,25-дигидроксивитамином D3 и РВD повышает эффективность кишечной абсорбции Са 2+ до 30–40%, т. е. в 2–4 раза, а фосфора — до 80%. Сходные механизмы действия D-гормона лежат в основе осуществляемой под его влиянием реабсорбции Са 2+ в почках.
В костях 1α,25(ОН)2D3 связывается с рецепторами на кость-формирующих клетках — остеобластах, вызывая повышение экспрессии ими лиганда рецептора активатора ядерного фактора кВ (RANKL). Рецептор-активатор ядерного фактора кВ (RANK), являющийся рецептором для RANKL, локализованным на преостеокластах, связывает RANKL, что вызывает быстрое созревание преостеокластов и их превращение в зрелые остеокласты. В процессах костного ремоделирования зрелые остеокласты резорбируют кость, что сопровождается выделением кальция и фосфора из минерального компонента (гидроксиапатита) и обеспечивает поддержание уровня кальция и фосфора в крови. В свою очередь, адекватный уровень кальция (Са 2+ ) и фосфора необходим для нормальной минерализации скелета [11–13].
Многочисленные исследования показали, что назначение препаратов кальция и/или витамина D способствует уменьшению потери костной ткани [14–19]. У женщин в поздней постменопаузе с низким употреблением пищевого кальция прием кальция предотвращает потерю костной ткани в позвоночнике [20, 21]. В свою очередь, назначение добавок кальция лицам старше 60 лет приводит к снижению потери костной массы в области бедра среди белых мужчин и женщин в возрасте моложе 72 лет [22]. Эффект назначения цитрата кальция на минеральную плотность кости (МПК) у женщин в раннем (до 5 лет) и среднем (от 5 до 10 лет) постменопаузальном периоде в течение двух лет проявлялся в виде прироста МПК в поясничном отделе на 1%, наряду со значимым снижением МПК на 2,4% в группе, получавшей плацебо [23]. Метаанализ 9 рандомизированных клинических исследований с общей выборкой более 50 тыс. человек, в 6 из которых сравнивалось комбинированное лечение витамином D (400 или 700–800 МЕ/сут) и кальцием с группами плацебо или без лечения, продемонстрировал достоверное снижение риска перелома бедра на 18% (RR 0,82 [95% ДИ 0,71–0,94], р = 0,0005) и риска внепозвоночных переломов на 12% (RR 0,88 [95% ДИ 0,78–0,99], р = 0,036) в группах, получавших комбинированную терапию, по сравнению с группами без добавок [24]. В исследованиях, где применялся витамин D в дозе 700–800 МЕ/сут, эффект на риск перелома бедра был выше, чем при приеме 400 МЕ (21% и 18% соответственно). Соответственно, в исследованиях, в которых пациенты получали только витамин D или плацебо (4 РКИ с общей численностью 9083 пациента), не было получено снижения риска внепозвоночных переломов как при применении дозы 400 МЕ (RR 1,14 [95% ДИ 0,87–1,49]), так при использовании 700–800 МЕ (RR 1,04 [95% ДИ 0,75–1,46]), что подтверждает ранее представленные данные о том, что витамин D без добавления кальция не снижает риск переломов [24].
Магний
Известно, что 60–65% магния находится именно в скелете и от обеспеченности костей магнием зависит обмен кальция и витамина D. Являясь структурным компонентом значительного числа ферментов, магний образует кристаллы с фосфатами, принимает участие в росте и стабилизации кристалла гидроксиапатита — структурной единицы минерального компонента костной ткани [25, 26]. Магний регулирует секрецию паратгормона (ПГ), повышает чувствительность клеток-мишеней к ПГ и витамину D, стимулирует действие кальцитонина. Длительное во времени нарушение соотношения Mg/Ca в сторону дефицита магния сопровождается замедлением обменных процессов в кости. Специальные магний-дефицитные диеты, сопровождающиеся уменьшением сывороточной концентрации магния, способствуют системной потере костной массы, снижению толщины надкостницы, характерным изменениям провоспалительных маркеров и маркеров резорбции кости. Уже по истечении достаточно короткого срока (4 недели), магний-дефицитная диета приводит к значимому снижению содержания минеральных веществ кости (р
М. И. Шупина, кандидат медицинских наук
Г. И. Нечаева 1 , доктор медицинских наук, профессор
Д. В. Шупин
Е. В. Надей
А. А. Семенкин, доктор медицинских наук, профессор
ГБОУ ВПО ОмГМУ МЗ РФ, Омск
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом