
ХРЯЩЕВАЯ ТКАНЬ [textus cartilagineus (LNH)] — разновидность соединительной ткани, выполняет опорную функцию.
Xрящевая ткань входит в состав скелета (см.) в виде хрящевых покрытий суставных поверхностей костей (суставной хрящ), хряща межпозвоночных дисков, реберных хрящей, а также формирует внескелетные опорные структуры (хрящи гортани, трахеи, бронхов, хрящевую часть евстахиевой трубы, хрящевые пластинки ушной раковины, носа и др.).
В эмбриогенезе хрящевая ткань образуется из мезенхимы (см.). Предшественниками хондроцитов являются мало-дифференцированные прохондробласты и хондробласты. Они составляют основную массу закладки хрящевой ткани в процессе гистогенеза, а в дальнейшем присутствуют в надхрящнице. На ранних этапах внутриутробного развития почти весь скелет высших позвоночных и человека является хрящевым. В этот период хрящевая ткань составляет до 45% веса (массы) тела. В процессе антенатального и раннего постнатального развития хрящевая ткань в основном замещается костной тканью (см. Кость), в результате чего у взрослого человека масса всех хрящевых образований не превышает 2% массы тела.
Все хрящевые образования за исключением суставного хряща покрыты надхрящницей, состоящей из плотной волокнистой соединительной ткани, богатой сосудами. Надхрящница обеспечивает рост и питание хряща. Кроме того, питание суставного хряща осуществляется при активном участии синовиальной жидкости (см.), находящейся в суставной полости.
Хрящевая ткань состоит из хондроцитов (хрящевых клеток) и хрящевого матрикса. Хондроциты представляют собой крупные клетки овальной или округлой формы с небольшими отростками. В зависимости от степени зрелости различают хондроциты нескольких типов. Хондроцит I типа — молодая активная клетка с высокими показателями синтеза ДНК, обладающая способностью к митозу (см.). Зрелые хондроциты II и III типа содержат в цитоплазме хорошо развитую эндоплазматиче-скую сеть и комплекс Гольджи, активно продуцируют и секретируют коллаген, гликопротеиды, протеогликаны. Для них характерен амитотический тип деления (см. Амитоз).
В зрелом хряще присутствуют также пузырчатые сильно вакуолизированные разрушающиеся клетки, окончившие свой жизненный цикл. Хондроциты находятся в полостях (лакунах) матрикса изолированно или группами, образовавшимися в результате деления одной клетки (изогенные группы). Стенки лакуны представляют собой плотный волокнистый коллагеновый каркас (так наз. перицеллюлярную корзину), который защищает клетки от механических воздействий. Внутри лакуны хондроциты окружены тонкофибриллярным богатым водой основным веществом.
В хрящевом матриксе различают волокна и основное вещество. Волокнистыми компонентами хрящевого матрикса являются коллаген II типа, эластин, белки неколлагеновой природы, гликопротеиды, протеогликаны. Макромолекулы коллагена образуют волокнистые структуры при взаимодействии с гликопротеидами и протеогликанами.
Основное вещество состоит из протеогликанов и гликопротеидов и не является аморфным. Выявлена строгая ориентация, упорядоченность в расположении макромолекул и их агрегатов; векторами ориентации являются как направление волокон коллагена, так и расположение хондроцитов.
Хрящевой ткани свойственна четкая упорядоченность во взаимном расположении клеток и матрикса. В ней принято различать территориальные и межтерриториальные участки. Территориальные участки образованы изогенными группами клеток, окруженных основным веществом и ограниченных волокнистым каркасом циркулярно расположенных коллагеновых волокон. Межтерриториальные участки представлены пучками волокон с прослойками основного вещества, ориентированных соответственно вектору силовых линий распределения нагрузки.
В зависимости от преобладания тех или иных волокнистых компонентов и степени маскировки их гомогенным основным веществом принято различать гиалиновый, волокнистый и эластический хрящи. Наиболее часто в организме встречается гиалиновый хрящ. Гиалиновыми являются суставные и реберные хрящи, а также хрящи носа, гортани (щитовидный и перстневидный), эпифизарный хрящ длинных трубчатых костей, хрящи трахеи и бронхов. Нативный гиалиновый хрящ — плотный, упругий, жемчужно-белый (стекловидный), что связано со значительным содержанием в нем гомогенного основного вещества, богатого протеогликанами, при удалении которых выявляется волокнистый коллагеновый каркас.
Для волокнистого хряща характерно наличие выраженных пучков коллагеновых волокон, а также гетерогенность клеток (наряду с хондроцитами в нем присутствуют и фибробласты). Из волокнистого хряща построены межпозвоночные диски, непрерывные соединения (синхондрозы), а также участки сухожилий и связок в месте их прикрепления.
Эластический хрящ обнаруживается в ушной раковине, надгортаннике, рожковидных и черпаловидных хрящах гортани. Он отличается большим содержанием в матриксе эластических волокон и не подвергается обызвествлению.
Своеобразной разновидностью хрящевой ткани является хондроидная ткань стромы сердца, сохраняющаяся в отдельных участках фиброзных колец у взрослых.
Читайте также: Что значит ткань футер двухнитка
Регенерация хрящевой ткани осуществляется за счет малодифференцированных клеток надхрящницы, а также, по-видимому, благодаря способности хондроцитов при определенных условиях к митотическому делению.
Биохимия хрящевой ткан и химический состав хрящевой ткани в связи с бедностью клетками практически полностью определяется составом ее матрикса, или межклеточного вещества (см.). Хрящевая ткань богата водой (более 70%); сухой остаток составляет около 30%, в нем содержится примерно 50% коллагена (см.), причем специфическим для хрящевой ткани является коллаген II типа, молекулы которого состоят из трех одинаковых полипептидных альфа-цепей. Кроме того, в хрящевой ткани содержится несколько своеобразных, так называемых минорных, коллагенов. В нормальном гиалиновом хряще коллаген II типа составляет основную массу коллагена, в эластическом и фиброзном хрящах наряду с коллагеном II типа присутствует также коллаген I типа. В суставных хрящах концентрация коллагена наиболее высока в поверхностном слое.
Другим компонентом хрящевой ткани являются гликозаминогликаны (см. Мукополисахариды), суммарное содержание которых в эмбриональном периоде достигает 25% сухого остатка, затем постепенно снижается и в пожилом возрасте составляет 14%. Сульфатированные гликозаминогликаны — хондроитинсульфаты (см. Хондроитинсерные кислоты) и кератансульфат — присоединены к так наз. стержневому белку и образуют макромолекулы протеогликанов (протеинполисахаридов, хондромукопротеинов) массой 1 000 000 — 3 000 000. Особенностью хрящевой ткани является то, что протеогликаны соединяются с помощью гиалуроновой кислоты (см. Гиалуроновые кислоты) в агрегаты массой до 50 000 000 — 100 000 000. Агрегаты протеогликанов удерживают в связанном состоянии основную часть содержащихся в хрящевой ткани воды и растворов электролитов, благодаря осмотическому эффекту способствуют поддержанию коллагенового каркаса в расправленном состоянии и обеспечивают диффузию веществ в хрящевую ткань, не содержащую кровеносных сосудов.
Белки неколлагеновой природы составляют 10—20% сухого остатка, в том числе белки, связанные с гликозаминогликанами 7 — 13%, структурные гликопротеиды (см.) и липопротеиды (см.) 3 — 7%, липиды (см.) и дезоксирибонуклеиновые кислоты (см.) 1,3—1,8%. Клетки и матрикс хрящевой ткани содержат также гликоген (см.), а хрящевая ткань эпифизарных отделов костей — фосфорно-кальциевые соли.
Обмен веществ в хрящевой ткани в связи с отсутствием васкуляризации осуществляется хондроцитами (благодаря физическим свойствам и специфическому строению матрикса) и обеспечивается энергией за счет гликолиза (см.), протекающего преимущественно по анаэробному типу. Обмен веществ весьма интенсивен в период роста, особенно в эпифизарных хрящах, но затем резко замедляется, и зрелая хрящевая ткань характеризуется выраженной метаболической инертностью. Xрящевая ткань обладает способностью к обратимой деформации в условиях значительных механических нагрузок, а также слабой иммунологической реактивностью в связи со способностью гидратированного матрикса задерживать и изолировать антигены.
В процессе старения организма в хрящевой ткани уменьшается концентрация протеогликанов, а следовательно, и стелень гидратированности матрикса.
В хондроцитах накапливаются гликоген и липиды, уменьшаются размеры комплекса Гольджи (см. Гольджи комплекс) и эндоплазматической сети (см. Эндоплазматический ретикулум), а также число митохондрий (см.). Клетки вакуолизируются и гибнут, а лакуны заполняются основным веществом. В матриксе откладываются соли кальция и уменьшается содержание воды, что ведет к потере хрящом эластодинамических свойств.
В патологических условиях обмен веществ в хрящевой ткани нарушается: повышается активность протеолитических ферментов, интенсифицируются катаболические и биосинтетические процессы, происходит нарушение структуры и агрегации протеогликанов, появляются несвойственные хрящевой ткани коллагены, отмечаются отложение пигментов и избыток липидов.
Библиогр.: Жаденов И. И. и Пастель В. Б. Обменные процессы в суставном хряще в норме (возрастной аспект) и при патологии (остеоартроз), Ортоп. и травмат., № 3, с. 65, 1982, библиогр.; Многотомное руководство по ортопедии и травматологии, под ред. Н. П. Новаченко, т. 1, с. 427, 606, М., 1967; Многотомное руководство по патологической анатомии, под ред. А. И. Струкова, т. 5, с. 234, 433, М., 1959; Павлова В. Н. Синовиальная среда суставов, с. 155, М., 1980; Рейнберг С. А. Рентгенодиагностика заболеваний костей и суставов, кн. 1, М., 1964; Слуцкий Л. И. Биохимия нормальной и патологически измененной соединительной ткани, Л., 1969; Тагер И. Л. Рентгенодиагностика заболеваний позвоночника, с. 101, М., 1983; Франке К. Спортивная травматология, пер. с нем., с. 74, М., 1981; Хэм А. и Кормак Д. Гистология, пер. с англ., т. 3, М., 1983; KneseK. -H. Stiitzgewebe und Skelett-system, В. u. а., 1979; Thompson R. С. a. Robinson H. J. Articular cartilage matrix metabolism, J. Bone Jt. Surg. v. 63-A, p. 327, 1981.
Читайте также: Физические свойства ткани характеризуют
В. H. Павлова (ан., гист., эмбр.), Л. И. Слуцкий (биохим.).
Минорный коллаген хрящевой ткани появляющийся при патологическом обызвествление хряща тип
от 400 до 1000 кгс/см2) в зависимости от времени, а температуры от 60 °C до 70 °С возрастает на 20 %. Наибольшая степень гидролиза наблюдается при температуре протеолиза, равной 42 °С. В условиях действия химопсина получены пептиды средней глубины гидролиза. Длина гидролизованых пептидов коллагена соответствует 10-30 аминокислотным остаткам, а их молекулярные массы изменяются от 1000 до 6000 Да.

Все виды хрящевой ткани состоят из клеток и внеклеточного матрикса (ВКМ). ВКМ образован из коллагеновых фибрилл и протеогликанов. Механическая прочность матрикса хрящевой соединительной ткани определяется жесткой структурой молекул и фибрилл коллагена [1]. В хрящах преобладают молекулы коллагена типа II, а фибриллы формируются из комплексов коллагенов типа II и типов I, III, IX, X, XI, XII, XIV. Минорные фибриллы коллагена типа VI связывают основные фибриллы между собой. Разрушения матрикса хрящевой ткани возникают при остеоартрозе, ревматоидном артрите, полихондрии. Во всех суставах изменения при болезнях начинаются прежде всего в гиалиновых хрящах. Происходит истончение фибрилл коллагена, их разволокнение и потеря эластичности. Ослабление структуры фибрилл в суставах наблюдается также при наследственном заболевании гипермобильности суставов, которое сопровождается истончением кожи. Для образования прочных коллагеновых фибрилл и сохранения целостности ткани необходим специфический состав компонентов.
Формирование коллагеновых фибрилл хрящевой соединительной ткани в организме человека происходит из тех аминокислот, белков, гликозаминогликанов, витаминов, микро- и макроэлементов, которые поступают преимущественно с пищей. Полный набор коллагенов разного типа можно получить из аналогичных тканей здоровых сельскохозяйственных животных. Коллагеновые белки, предварительно денатурированные температурой и поступающие с пищей, под действием ферментов желудочно-кишечного тракта могут расщепляться до пептидов и аминокислот. Аминокислоты всасываются в кровь, а затем поступают в клетки матрикса соединительных тканей. Биодоступность коллагена и гликозаминогликанов для усвоения организмом является более высокой после ферментативного гидролиза in vitro.
Параметры гомогенизации гиалиновых хрящей трахей крупного рогатого скота
t (время гомогенизации), мин.
Средний диаметр частиц, унимодальный анализ (нм)
Во многих странах коллагеновые гидролизаты применяют в медицине с целью профилактики и лечения болезней человека, укрепления структуры фибрилл 3. Например, клинические испытания показали, что состояние больных остеоартрозом облегчается в результате совместного действия гидролизованного коллагена и витамина С [2]. Коллагеновые гидролизаты после приёма в течение длительного времени улучшили функционирование коленного сустава у пациентов не только с облегчённой формой остеоартроза, но и с его сложной формой [4].
Целью данной работы является исследование протеолитического расщепления коллагенов хрящевой ткани и получение пептидов регулируемых размеров.
Материалы и методы исследования
Гиалиновые хрящи были выделены из трахей крупного рогатого скота. Биокаталитический метод способствует сохранению структуры ингредиентов матрикса. Для гидролиза белков, содержащихся в гомогенате хрящей, был использован комплекс ферментов (химопсин, панкреатин), действующий в слабощелочной среде. Был исследован протеолиз коллагеновых белков хрящевой ткани в зависимости от температуры, кислотности, концентрации ферментных препаратов и продолжительности процесса. Степень гидролиза (DH) является характеристикой глубины гидролиза. Индекс DH коррелирует с распределением длин молекулярных цепей и с молекулярной массой пептидов. Для оценок качественного состава гидролизатов мы использовали спектрофотометрию, ЯМР-спектроскопию и МАЛДИ масс-спектроскопию.
Результаты исследования и их обсуждение
Гиалиновые хрящи крупного рогатого скота были получены сразу же после забоя животных. Хрящи, очищенные от пленок, сначала были измельчены вручную до частиц размером 3–4 мм, затем были диспергированы и гомогенизированы в водной суспензии до размеров частиц размером 3–4 мкм в механическом гомогенизаторе тканей типа миксера Braun MQ 520. Затем были получены более мелкие размеры частиц менее 700-800 нм в гомогенизаторе высокого давления (ГВД) при давлении 20–60 МПа (
Читайте также: Как убрать жвачку с ткани быстро
200-600 кгс/см2). ГВД «Донор-3» был разработан В.В. Капцовым в ИБК РАН, г. Пущино на базе ГВД «Донор-1». Еще меньше размеры частиц 300–400 нм были получены при дальнейшей гомогенизации суспензии хрящевых частиц в условиях последовательного повышения давления и температуры в течение 20–60 мин («тонкая гомогенизация»). Были исследованы образцы (A, B, C, E, F) со следующими параметрами «тонкой» гомогенизации (табл. 1).
Анализ размеров хрящевых частиц в водной суспензии проводился на фотонном допплеровском анализаторе частиц фирмы Beckman-Coulter N5 (США) двумя методами анализа. В основном измерялся средний диаметр хрящевых частиц при допущении их сферической формы по унимодальному анализу – определение среднего диаметра сферических частиц при нормальном логарифмическом распределении частиц без анализа возможности выявления нескольких пиков распределения частиц в водной суспензии гомогенизированных хрящей. На рис. 1 приведён график распределения частиц образца F в гомогенизированной хрящевой суспензии при унимодальном анализе.
Результаты унимодального распределения при угле 90 °
Средний диаметр частицы (нм)
Коэффициент диффузии (кв. м/сек.)
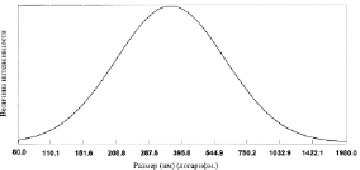
Рис. 1. Средний диаметр частиц гомогенизированной хрящевой суспензии (393,6 нм) измеренный на фотонном допплеровском анализаторе частиц фирмы Beckman-Coulter N5 (США) по унимодальному анализу (образец F)
Анализ распределения частиц по диаметрам при угле 90 °
Диапазон распределения размеров (нм)
Среднее стандартное отклонение
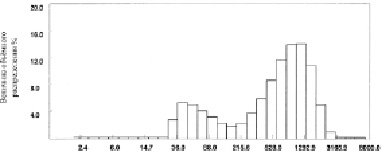
Рис. 2. Распределение частиц по диаметрам гомогенизированной хрящевой суспензии при измерении на фотонном допплеровском анализаторе частиц фирмы Beckman-Coulter N5 (США) по SDP анализу (образец F)
На рис. 2 приведён график распределения частиц образца F в гомогенизированной хрящевой суспензии при проведении SDP (size distribution processor) анализа, т.е. при измерении размеров частиц с анализом на мультимодальное (многопиковое) распределение. Сравнивая распределение хрящевых частиц в суспензии у различных образцов, следует отметить их приблизительно одинаковый характер для всех образцов «тонкой» гомогенизации.
Ферментативный гидролиз гиалиновых хрящей
Ферментативный гидролиз является предпочтительным для получения продуктов расщепления биополимеров соединительных тканей по сравнению с другими методами. В результате ферментативного гидролиза не нарушается структура аминокислот и их состав. Гидролизаты содержат комплекс аминокислот и пептидов, которые не требуют больших затрат энергии в процессе пищеварения и усвоения. Гидролиз гиалиновых хрящей проводили последовательными процедурами. Как правило, для гидролиза белков, содержащихся в соединительных тканях, используют не отдельные протеазы, а комплексы ферментов [5]. Эти ферменты обладают специфичным действием, разрывая определенные связи в белках. Мы изучали гидролиз белков хрящевой ткани, применяя химопсин и панкреатин. Оптимум рН для этих ферментов находится в интервале 7,5 – 9,0, т.е. в области рН, которая ниже рК (10,5) молекулы коллагена. Сравнительный анализ действия этих ферментов при одинаковых условиях рН (8,0), температуры (42 °С), концентрации фермента (1 %) и времени (4 час.) показал, что параметр DH больше в гидролизатах, полученных после действия панкреатина. Однако достоверной разницы между образцами мы не обнаружили. Относительно низкую степень гидролиза можно повысить как гомогенизацией хрящевой ткани, так и параметрами протеолиза.
Наши исследования были направлены на активизацию ферментативного гидролиза гомогената хрящей под действием химопсина. В процессе получения гомогенизированных образцов мы варьировали значения давления, температуры и времени, которые соответствовали параметрам табл. 1. Было проверено влияние химопсина на гомогенаты гиалиновых хрящей, полученные в разных условиях денатурирующего действия давления и температуры (образцы А, В, С, Е, F). Известно, что ферменты химотрипсин и трипсин, содержащиеся в химопсине, эффективно расщепляют полипептидные цепи тройной спирали молекулы коллагена после её денатурации.
Гидролиз проводили в K-Na фосфатном буфере при фиксированных значениях рН (8,0) и температуры (42 °С) в зависимости от концентрации фермента и времени. Гидролиз продолжался в течение 8 часов, но максимальная степень гидролиза наблюдалась после 5–6 часов. Данные по степени гидролиза в разных гомогенатах хрящевой ткани под действием химопсина приведены в табл. 2.
Степень гидролиза ферментативных гидролизатов, полученных под влиянием химопсина на разные гомогенаты хрящевой ткани
- Правообладателям
- Политика конфиденциальности