
Описан состав, строение и функции миофибриллы. При силовой тренировке наблюдается миофибриллярная гипертрофия – увеличение количества и объема миофибрилл. У детей гипертрофия миофибрилл происходит за счет их роста в длину, при силовой тренировке – в толщину.
Состав, строение и функции миофибриллы
Общая характеристика
Для того чтобы понять, какие механизмы лежат в основе гипертрофии мышц, нужно рассмотреть состав, строение (структуру) и функции миофибриллы.
Миофибриллы представляют собой органеллы специального назначения мышечного волокна. Это – тонкие белковые нити, расположенные вдоль мышечного волокна параллельно друг другу (рис.1). Миофибриллы, в отличие от других компонентов мышечного волокна, не имеет оболочки. Роль оболочки играет саркоплазматический ретикулум, который окружает каждую миофибриллу в виде «муфточки». Миофибриллы идут от одного конца мышечного волокна до другого, их длина соответствует длине волокна.
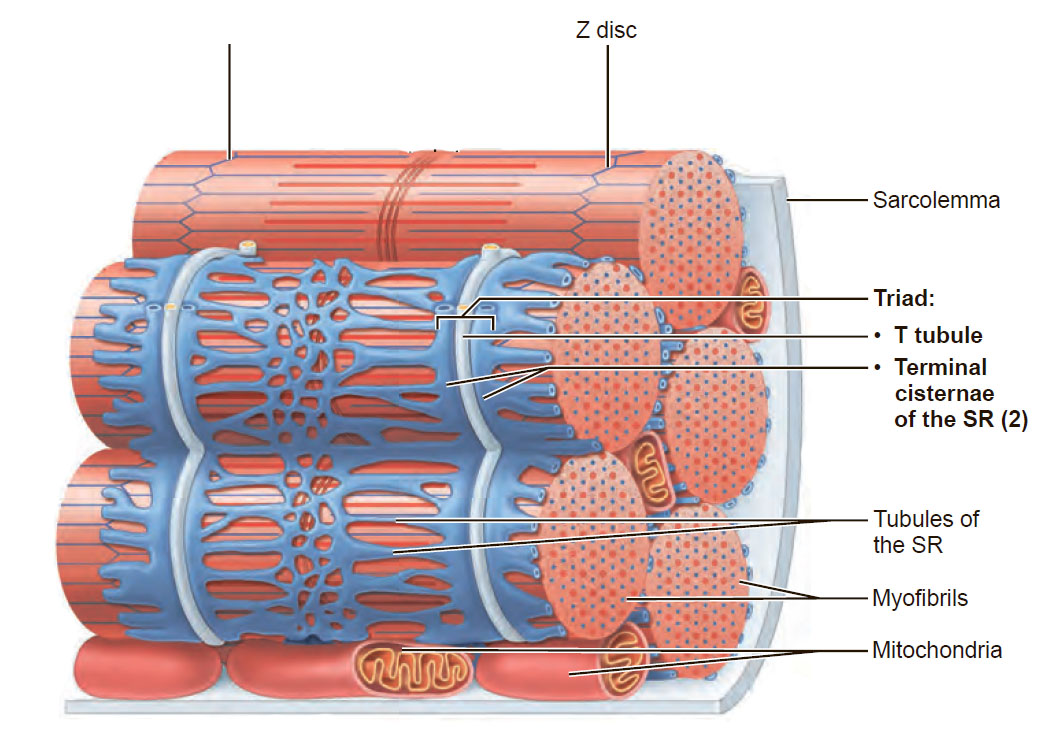
Рис.1. Миофибриллы мышечного волокна, окруженные саркоплазматическим ретикулумом
Функции миофибриллы
Миофибриллы – основные сократительные элементы мышечного волокна, поэтому их основная функция — укорочение под воздействием нервного импульса. Вследствие этого мышца развивает определенную силу.
Состав миофибриллы
Миофибриллы состоят из элементов, имеющих цилиндрическую форму – саркомеров, которые расположены последовательно, друг за другом вдоль миофибриллы (рис.2). Друг от друга саркомеры отделены Z-дисками (в плоскости – Z-линии). Миофибриллу можно сравнить со стеблем бамбука, длинные секции которого соединяются друг с другом толстыми дисками. Длина одного саркомера в среднем равна 2,5 мкм, поэтому в одной миофибрилле длиной 5 см находится до 20000 саркомеров.
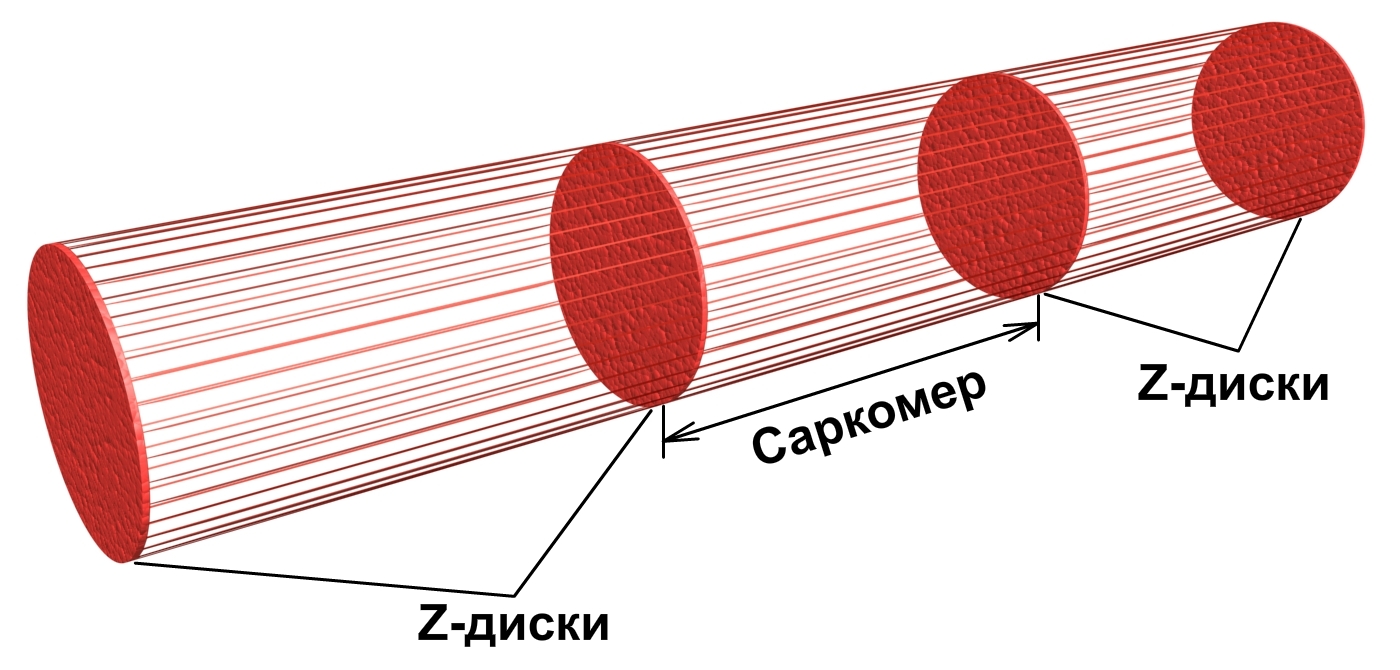
Рис.2. Миофибрилла состоит из саркомеров
Расположение в мышечном волокне
В мышечном волокне может содержаться от нескольких десятков до нескольких тысяч миофибрилл. Каждая миофибрилла в мышечном волокне «привязана» к соседней посредством белковых соединений, которые называются промежуточными филаментами. Все периферические миофибриллы имеют связь с оболочкой мышечного волокна (сарколеммой) посредством белковых структур, которые называются костамерами.
Рис. 3. Связь периферических миофибрилл с оболочкой мышечного волокна (сарколеммой) посредством белковых структур, которые называются костамерами.
Влияние силовой тренировки на объем и количество миофибрилл
Доказано, что под влиянием силовой тренировки в мышечном волокне увеличивается количество миофибрилл и их объем. Это явление называется миофибриллярной гипертрофией.
Увеличение количества миофибрилл называется гиперплазией миофибрилл. Увеличение объема миофибрилл называется гипертрофией миофибрилл.
У взрослых объем миофибрилл возрастает за счет изменения площади поперечного сечения миофибриллы (она становится толще). Ученый Голдспинк еще в 1970 году доказал, что при достижении миофибриллы больших размеров, она расщепляется на две миофибриллы.
Гипертрофия миофибрилл у детей
У детей происходит увеличение длины мышц, соответственно «растут» в длину и миофибриллы. Это происходит за счет добавления саркомеров на концах миофибрилл. Таким образом увеличивается объем миофибрилл, то есть происходит их гипертрофия.
Неиспользование
Если конечность травмирована, например, наложен гипс, и фиксирована длина мышцы, через несколько часов длина мышцы начинает уменьшаться. Длина миофибрилл также уменьшается. Это происходит за счет разрушения саркомеров, расположенных на краях миофибрилл. Кроме того, происходит уменьшение толщины миофибрилл и их количества.
СПЕЦИАЛЬНЫЕ ОРГАНОИДЫ И ВКЛЮЧЕНИЯ
Описание: С помощью ресничек и жгутиков клетки могут передвигаться в жидкой среде так как эти органоиды способны совершать ритмические движения. Если на поверхности клетки имеется большое количество волосковидных выростов небольшой длины то их называют ресничками если же таких выростов мало и длина их значительная то они называются жгутиками. Клетки высших растений и высших грибов а также споровики не имеют ресничек и жгутиков даже у мужских половых клеток. Миофибриллы Миофибриллы представляют собой особые дифференцированные сократимые элементы.
Поделитесь работой в социальных сетях
Если эта работа Вам не подошла внизу страницы есть список похожих работ. Так же Вы можете воспользоваться кнопкой поиск
СПЕЦИАЛЬНЫЕ ОРГАНОИДЫ И ВКЛЮЧЕНИЯ
Органоиды специального назначения
Органоиды специального назначения содержатся во многих животных и растительных клетках. От общих органоидов они отличаются тем, что характерны только для определенных высоко дифференцированных клеток и выполняют строго определенную функцию, характерную для этих клеток.
Классификация органоидов специального назначения:
1. Органоиды движения: реснички, жгутики, миофибриллы.
2. Опорные структуры: тонофибриллы.
3. Органоиды, участвующие в передаче возбуждения: нейрофибриллы.
4. Органоиды, воспринимающие внешние раздражения: фоторецепторы, статорецепторы, фонорецепторы.
5. Органоиды поверхности клеток: микроворсинки, кутикула.
6. Органоиды защиты и нападения у одноклеточных: трихоцисты у инфузорий; коноид, роптрии у представителей класса Споровиков.
Рассмотрим более подробно основные из этих органоидов.
Реснички и жгутики это нитевидные или волосковидные выросты свободной поверхности клеток. С помощью ресничек и жгутиков клетки могут передвигаться в жидкой среде, так как эти органоиды способны совершать ритмические движения. Если же реснички и жгутики имеются у прикрепленных к какому-либо субстрату клеток, то они вызывают движение окружающей жидкости.
Различий в тонкой организации этих структур нет. Если на поверхности клетки имеется большое количество волосковидных выростов небольшой длины, то их называют ресничками , если же таких выростов мало и длина их значительная, то они называются жгутиками .
У животных реснички и жгутики встречаются: а) в клетках ресничного эпителия (эпителий трахеи, некоторых отделов полового тракта); б) у сперматозоидов (у нематод и десятиногих раков спермии не имеют жгута); в) у простейших (жгутиконосцы, инфузории, корненожки). В мире растений они имеются у подвижных зооспор водорослей, мхов, папоротников, низших грибов, миксомицетов. Клетки высших растений и высших грибов, а также споровики не имеют ресничек и жгутиков даже у мужских половых клеток.
Читайте также: Как сделать зеленые листья из ткани
Толщина ресничек и жгутиков составляет около 200 нм (0,2 мкм). Поскольку принципиальных различий в строении ресничек и жгутиков нет, рассмотрим ультраструктуру этих образований на примере реснички. Снаружи ресничка покрыта цитоплазматической мембраной. Внутри нее расположена аксонема (или осевой цилиндр), состоящая из микротрубочек. Нижняя проксимальная часть реснички, базальное тельце , погружена в цитоплазму. Диаметры аксонемы и базального тельца одинаковы.
Базальное тельце по своей структуре совершенно сходно с центриолью и состоит из 9 триплетов микротрубочек. Аксонема в своем составе, в отличие от базального тельца, имеет 9 пар (дублетов) микротрубочек, образующих внешнюю стенку цилиндра аксонемы. Дублеты микротрубочек слегка повернуты (около 10 0 ) по отношению к радиусу аксонемы. Кроме периферических дублетов микротрубочек в центре аксонемы располагается пара центральных микротрубочек. Эти две центральные микротрубочки, в отличие от периферических, не доходят до базальных телец. Поскольку в базальных тельцах содержится сократимый белок типа актомиозина, периферические микротрубочки выполняют двигательную функцию, а центральные только опорную.
В основании ресничек и жгутиков часто встречаются корешки или кинетодесмы , представляющие собой пучки тонких (6 нм) фибрилл, обладающих поперечных исчерченностью. Часто такие исчерченные кинетодесмы простираются от базальных телец в глубь цитоплазмы в направлении к ядру. Роль этих структур еще недостаточно выяснена.
Отклонения от вышеизложенного плана строения встречаются редко, но у некоторых клеток, например, в жгутиках сперматозоидов и некоторых жгутиконосцев, обнаружены 9 дополнительных фибрилл, расположенных между центральными и периферическими микротрубочками. Эти дополнительные фибриллы соединены с трубочками аксонемы с помощью очень тонких волокон.
Миофибриллы представляют собой особые дифференцированные сократимые элементы клетки, за счет которых происходят сложные и совершенные движения мышц. Различают два типа миофибрилл: гладкие и поперечнополосатые. Оба типа миофибрилл широко распространены у многоклеточных животных и у простейших.
Поперечнополосатые миофибриллы широко известны в составе соматической и сердечной мускулатуры членистоногих и хордовых животных. Гладкие миофибриллы типичны для мускулатуры внутренних органов позвоночных и для соматических мышц многих низших беспозвоночных.
Строение миофибрилл наиболее подробно изучено в поперечно-полосатых мышечных волокнах. Миофибрилла имеет толщину 0,5 мкм и длину, которая равна от 10-20 мкм до нескольких миллиметров и даже сантиметров. В световой микроскоп видно, что пучки миофибрилл окрашиваются неравномерно: через равные промежутки длины в них видно чередование темных и светлых участков. Темные участки имеют двойное лучепреломление и называются анизотропными дисками (А-диски) . Светлые участки двойного лучепреломления не обнаруживают и называются изотропными дисками ( I -диски) .
Каждый А-диск разделяется на две половины менее плотной, чем остальные его участки, полосой, называемой Н-зоной (полоска Ханзена). Посередине каждого I -диска проходит темная линия, называемая Z -линией (телофрагма) . Участок миофибриллы между двумя Z -линиями называется саркомером. Он является единицей строения и функционирования миофибриллы.
Подробности строения саркомера были получены только при изучении миофибрилл в электронном микроскопе. Каждая миофибрилла состоит из пучка очень тонких нитей миофиламентов. Различают два типа миофиламентов: толстые и тонкие. Тонкие миофиламенты имеют диаметр около 7 нм и длину около 1 мкм; они состоят в основном из белка актина. Они располагаются в пределах I -диска и заходят в А-диск до Н-зоны. Толстые миофиламенты длиной до 1,5 мкм и толщиной около 15 нм состоят из белка миозина; они расположены только в пределах А-диска. В тонких миофиламентах кроме актина находятся также белки тропомиозин и тропонин. Z -линии имеют в своем составе белок α-актинин и десмин.
Ни актин, ни миозин по отдельности не обладают сократительной способностью. Актин, белок с молекулярным весом 43,5 тысяч, является глобулярным белком размером около 3 нм. В присутствии АТФ и некоторых белковых факторов он способен к агрегации в виде нитчатых структур толщиной до 7 нм. Такие актиновые фибриллы состоят из двух спиралей, обвивающих друг друга. Миозин, входящий в состав толстых нитей, очень крупный белок (мол. вес 470 тысяч), состоящий из шести цепей: двух длинных, спирально обвивающихся одна вокруг другой, и четырех коротких, которые связываются с концами длинных цепей и образуют глобулярные «головки». Последние обладают АТФ-азной активностью, могут реагировать с фибриллярным актином, образуя актомиозиновый комплекс, способный к сокращению.
Актиновые миофиламенты связаны на одном конце с Z -линией, которая состоит из ветвящихся молекул белка α-актинина, образующих фибриллярную сеть, идущую поперек миофибриллы. С двух сторон к Z -линии прикрепляются концы актиновых нитей соседних саркомеров. Функция Z -линий заключается как бы в связывании соседних саркомеров друг с другом; Z -линии не являются сократимыми структурами.
Механизм мышечного сокращения заключается в одновременном укорачивании всех саркомеров по всей длине миофибриллы. Г. Хаксли показал, что в основе сокращения лежит перемещение относительно друг друга толстых и тонких нитей. При этом толстые миозиновые нити как бы входят в пространство между актиновыми нитями, приближая друг к другу Z -линии. Эта модель скользящих нитей может объяснить не только сокращение поперечнополосатых мышц, но и любых сократимых структур.
В гладких мышечных клетках также имеются актиновые и миозиновые нити, но они не так правильно расположены, как в исчерченных мышцах. Здесь нет саркомеров, а просто среди пучков актиновых миофиламентов без особого порядка располагаются миозиновые молекулы.
Читайте также: Ткани у марго ш энтузиастов 31 стр 38
Тонофибриллы характерны для клеток одноклеточных организмов и для эпителиальных клеток многоклеточных животных. Электронно-микроскопическое исследование показало, что они состоят из пучка тонофиламентов тончайших нитей с диаметром 6-15 нм. В одном пучке может быть от 3 до нескольких сотен тонофиламентов.
Тонофибриллы располагаются пучками в клетке в разных направлениях, прикрепляются либо к десмосомам, либо к любому участку цитоплазматической мембраны и никогда не переходят из одной клетки в другую.
Тонофибриллы выполняют в клетке опорную функцию.
Нейрофибриллы открыты в 1855 г. Ф.В. Овсянниковым. Они характерны для нервных клеток (нейронов). Состоят из более тонких нитей нейрофиламентов .
В теле нейрона нейрофибриллы расположены беспорядочно, а в отростках образуют пучок параллельно длине отростка. Из этого правила имеется всего лишь два исключения: параллельное, упорядоченное расположение нейрофибрилл в теле нейрона впервые обнаружено у бешеных животных, а затем у животных, которые впадают в спячку.
Открытие нейрофибрилл привело к возникновению нейрофибриллярной теории проведения нервного возбуждения. Сторонники этой теории считали, что нейрофибриллы являются беспрерывным проводящим элементом нервной системы. Однако в дальнейшем было установлено, что нейрофибриллы не переходят из одного нейрона в другой. В настоящее время мы придерживаемся нейронной теории , согласно которой в проведении нервного импульса основная роль принадлежит плазмалемме нейрона, а по нейрофибриллам из тела нейрона к его окончанию передаются вещества, участвующие в образовании нервных импульсов. А с одной клетки на другую возбуждение передается с помощью синапса (строение синапса описывалось ранее при рассмотрении коммуникационных межклеточных контактов). В синапсе возбуждение передается химическим путем с помощью медиатора.
Непостоянные включения в клетке
В отличие от органоидов, как общего, так и специального назначения, включения представляют собой непостоянные образования, то возникающие, то исчезающие в процессе жизнедеятельности клетки. Основное место локализации включений это цитоплазма, но они иногда встречаются и в ядре.
По своему характеру все включения это продукты клеточного метаболизма. По химическому составу и по выполняемым функциям они классифицируются следующим образом:
1. трофические (белковые, углеводные, жировые);
Белковые включения . Имеют форму зерен, гранул, дисков. Они могут присутствовать во всех клетках, но встречаются реже, чем жиры и углеводы. Примером белковых включений служит желток в яйцеклетках, алейроновые зерна в эндосперме семян. В этом случае белковые гранулы служат запасным питательным материалом для зародыша; в других клетках это трофический (строительный) материал для дальнейшего построения элементов клетки. Энергетическим запасом белковые включения могут служить в самом крайнем случае, когда углеводные и жировые запасы полностью израсходовались.
Углеводные включения также запасаются в твердом виде (глыбки, зерна разнообразных размеров и форм). У многоклеточных животных и простейших в цитоплазме клеток встречаются отложения гликогена. Обычно большие скопления гликогена сосредоточены в цитоплазме поперечнополосатых мышечных волокон, клетках печени, нейронах, у эндопаразитических организмов гельминтов и простейших. Гранулы гликогена могут располагаться не только в цитоплазме, но и в ядрах многих клеток, в частности, в ядрах клеток печени человека (более мелкие гранулы, чем в цитоплазме).
В клетках растений наиболее часто откладывается крахмал в виде зерен различной формы и размеров, причем форма крахмальных зерен специфична для каждого вида растений и для определенных тканей. Отложениями крахмала богата цитоплазма клубней картофеля, зерен злаков, бобовых растений и др. У низших растений встречаются другие полисахариды: парамилоид, крахмал красных водорослей.
Углеводные включения являются основным энергетическим запасом клетки. При распаде 1 г углевода выделяется 17,6 кДж энергии, которая накапливается в виде АТФ.
Жировые включения . Жиры в цитоплазме откладываются в виде мелких капель. Они встречаются как у животных, так и у растений. В одних клетках жировых включений очень мало и они постоянно используются самой клеткой в процессе обмена веществ, в других клетках они накапливаются в большом количестве, например, жировые клетки соединительной ткани, клетки эпителия печени рыб и амфибий. Большое количество жировых капель встречается и в цитоплазме многих видов простейших, например, инфузорий. Очень много жира содержится в семенах растений, причем количество его может доходить до 70% сухого веса семян (масличные культуры).
Процесс отложения жиров не связан с какими-либо органоидами клетки; они откладываются в основном веществе цитоплазмы. При определенных условиях жировые капли могут сливаться друг с другом, увеличиваясь в размерах, в конечном итоге гигантская жировая капля заполняет собой всю клетку, цитоплазма с ядром отмирают и клетка превращается в мешочек с жиром. Это явление называется жировое перерождение клетки . Этот процесс может носить патологический характер (например, при жировом перерождении печени, сердечной мышцы и т.д.) или являться естественным процессом в жизнедеятельности организма (например, клетки сальных желез, клетки подкожной жировой клетчатки китов, тюленей).
Жировые включения могут выполнять следующие функции:
1) являются долговременным энергетическим запасом клетки (при распаде 1 г жира выделяется 38,9 кДж энергии);
2) терморегуляция (например, у животных, обитающих в холодном климате слой жира в подкожной клетчатке достигает 1 м);
Читайте также: Аппликация из ткани вишня
3) амортизация при движении (например, прослойки жира на подошвах ног, на лапах у наземных животных, ладонях рук, вокруг внутренних органов);
4) запас питательных веществ у животных, впадающих в спячку (например, медведь, барсук, еж);
5) источник метаболической воды в организме у животных, обитающих в засушливых условиях (при распаде 1 кг жира образуется 1,1 кг воды).
Секреты это продукты анаболических реакций клетки, которые выполняют в организме различные жизненно важные функции.
Секреторные включения накапливаются в секреторных клетках в виде зерен, гранул, капель. Химическая природа их весьма разнообразна. Это могут быть белки, липиды, кетоны, спирты, соляная кислота и другие. В клетках многих растений встречаются и кристаллические включения, причем чаще всего это оксалаты кальция.
Функции секреторных включений:
1) гуморальная регуляция жизнедеятельности организма (гормоны в клетках желез внутренней секреции);
2) катализация процессов переваривания пищи (ферменты в клетках желез пищеварительного тракта);
3) передача возбуждения в синапсах (медиаторы в пресинаптических окончаниях нейронов);
4) питательные вещества для детенышей (молоко в млечных железах млекопитающих);
5) защитная функция (слизь у земноводных защищает кожу от пересыхания; яды, токсины у животных защищают от врагов и помогают умерщвлять добычу).
Из клеток секреты удаляются различными способами. По способу удаления секрета из клетки выделяют 3 типа секреции:
1) мерокриновая секрет удаляется через поры без повреждения клетки; такая клетка функционирует непрерывно (например, железы дна желудка);
2) апокриновая капли секрета отшнуровываются с частью цитоплазмы; такая клетка функционирует с перерывами, необходимыми для ее восстановления (например, слюнные железы, часть потовых)
3) голокриновая секрет заполняет клетку целиком, цитоплазма отмирает, клетка гибнет и превращается в мешочек с секретом; такая клетка функционирует всего один раз (например, сальные железы).
Экскреторные включения это продукты катаболических реакций, которые клеткой и организмом не используются, часто являются ядовитыми и должны удаляться. Экскреты могут накапливаться в жидком (капли) и в твердом (зерна, гранулы) состоянии.
Примерами экскреторных включений могут служить капли пота в клетках потовых желез, моча в клетках почечных канальцев. У многих беспозвоночных животных существуют специальные клетки нефроциты , которые функционируют как почки накопления. Они накапливают экскреты, а затем либо выносят их в кишечник или на поверхность тела, либо оставляют в составе своей цитоплазмы. Важную роль в обособлении ядовитых экскретов играет комплекс Гольджи. Примерами нефроцитов являются хлорагогенные клетки у кольчатых червей, перикардиальные клетки у моллюсков и насекомых, экскретофоры у ресничных червей и асцидий.
Пигментные включения могут существовать в виде гранул, зерен, изредка в виде капель. Основная их функция придание окраски растительным и животным клеткам и организму в целом. Но в ряде случаев пигментные включения выполняют более сложные функции. Рассмотрим в качестве примера некоторые пигменты животного и растительного мира.
1). Меланин пигмент коричневого цвета, расположен в клетках базального слоя кожи, придает окраску эпителию кожи и всем ее производным (волосы у человека, шерсть у животных, ногти, когти, перья у птиц, чешуи у рептилий), а также радужной оболочке глаза. У животных меланин создает различные виды защитной окраски, а у человека выполняет функцию защиты от ультрафиолетового излучения.
2). Липофусцин пигмент желтого цвета, гранулы которого накапливаются в процессе жизнедеятельности клеток и, особенно, по мере старения их, а также при разных дистрофических процессах («пигмент старения»).
3). Лютеин желтый пигмент, содержащийся в желтом теле беременности.
4). Ретинин характерный пигмент, входящий в состав зрительного пурпура сетчатки глаза.
5). Дыхательные пигменты животных:
гемоцианин пигмент, содержащий в своем составе медь; он может изменять свою окраску от синей (в окисленном состоянии) до бесцветной (в восстановленном состоянии); встречается у ракообразных, некоторых улиток, головоногих моллюсков (растворен в плазме крови или гемолимфе);
гемоэритрин пигмент, содержащий в своем составе железо; он может изменять свою окраску от красной (в окисленном состоянии) до бесцветной (в восстановленном состоянии); встречается у некоторых кольчатых червей (находится в клетках крови);
хлорокруорин пигмент, также содержащий в своем составе железо; он может изменять свою окраску от красной (в окисленном состоянии) до зеленой (в восстановленном состоянии); встречается у некоторых многощетинковых червей (растворен в плазме крови);
гемоглобин железосодержащий пигмент, меняет свою окраску от оранжево-красной (в окисленном состоянии) до пурпурно-красной (в восстановленном состоянии). Это наиболее широко распространенный в природе дыхательный пигмент, встречается у некоторых моллюсков (растворен в плазме крови), у некоторых кольчатых червей (в плазме или в клетках), у всех позвоночных животных (в эритроцитах крови).
Пигменты растительного мира:
1). Хлорофилл пигмент зеленого цвета, находится в гранах хлоропластов и участвует в процессе фотосинтеза.
2). Группа каротиноидов каротин (оранжевый), ксантофилл ( красный), ликопин (желтый); эти пигменты содержатся в хромопластах и обеспечивают окраску плодов, семян и других органов растений.
5). Фикобилины это пигменты низших растений; в состав сине-зеленых водорослей входит фикоциан (пигмент синего цвета), а в состав красных водорослей фикоэритрин (красный пигмент).
Изменение окраски клеток обусловлено перераспределением пигментов.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом