

По статистике, сердечно-сосудистые заболевания являются одной из основных причин инвалидности и смерти жителей большинства современных развитых стран [3]. На долю смертности от заболеваний сердечно-сосудистой системы в общем приходится до 60% от общего числа умерших.
Нередко для восстановления кровообращения в пораженных сосудах помимо медикаментозного лечения проводятся реконструктивные операции, и часто невозможно объективно оценить, какой тип оперативного вмешательства будет оптимальным для конкретного пациента, а также насколько близок будет кровоток в сосуде к нормальному после операции. Основная проблема при выполнении таких расчетов состоит в определении механических свойств стенок сосудов, параметров кровотока и др.
Еще одной важной проблемой при прогнозировании результатов лечения является скорость расчетов: как правило, большинство современных математических моделей требуют численного решения, причем во многих случаях вычисления получаются затратными по времени и требуют довольно мощные компьютеры [8]. При этом снижение времени расчетов путем упрощений может привести к неточности полученных результатов, что, безусловно, недопустимо [5].
Часто для численных расчетов применяют метод конечных элементов. Однако решение задач гемодинамики с помощью МКЭ требует больших затрат по времени [5].
Таким образом, актуальной является задача построения математической модели гемодинамики, которая бы достаточно полно описывала движение крови в кровеносных сосудах, учитывая взаимодействие жидкости со стенкой, и являлась легко адаптируемой под конкретного пациента.
Материалы и методы
Рассмотрим осесимметричное движение крови, которая принимается вязкой несжимаемой жидкостью, в круглом сосуде постоянного радиуса R. Движение происходит в цилиндрической системе координат (x, r, θ), причем ось x совпадает с осью симметрии потока [2]. Материал стенки считаем идеально упругим, изотропным.
Основная система уравнений динамики кровотока в гибких цилиндрических сосудах в таком случае будет иметь вид:
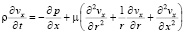
, (1)
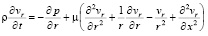
, (2)
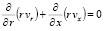
, (3)
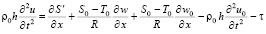
, (4)
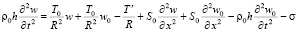
, (5)
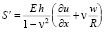
, (6)
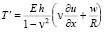
, (7)
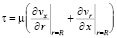 ,
, 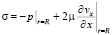 ,
,
где p – давление; ρ – плотность крови; μ – вязкость крови; vx – осевая компонента скорости крови; vr – радиальная компонента скорости крови; R – радиус сосуда; t – время; u, w – перемещения стенки в продольном и поперечном направлениях; Sʹ, Tʹ – силы натяжения в окружном и продольном направлениях соответственно; S0, T0 – начальные значения сил натяжения в окружном и продольном направлениях; E – модуль Юнга стенки; ν – коэффициент Пуассона; h – толщина стенки сосуда; ρ0 – массовая плотность материала стенки сосуда.
На стенке записываем условия кинематического контакта стенки сосуда с жидкостью:
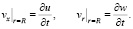
(8)
В случае моделирования гемодинамики крупных кровеносных сосудов в большинстве современных работ кровь полагается ньютоновской жидкостью [4, 7]. При этом в [6] показано, что разница значений (в случае крупных кровеносных сосудов), которые получаются для ньютоновской и неньютоновской жидкостей, не превышает 10%.
Уравнения (4)-(7) позволяют учесть податливость сосудистой стенки, а контактные условия (8) позволяют учесть взаимодействие стенки с потоком.
Такой подход к моделированию гемодинамики широко известен, однако основная система уравнений в этом случае не позволяет учесть конвективную составляющую ускорения частиц жидкости, а это, в свою очередь, для сосудистого русла с несколькими узлами бифуркации может оказать существенное влияние на результаты расчетов.
Основная система уравнений динамики вязкой несжимаемой жидкости в кровеносных сосудах с гибкими стенками в трехмерной постановке может быть записана в виде уравнений для направленных потоков.
Будем, как и раньше, полагать, что задача осесимметрична, а кровь является ньютоновской жидкостью.
Для направленных потоков жидкости в тонких трубках с упругими стенками на основе известного способа линеаризации возможен учет конвективного ускорения частиц жидкости в рамках линейной теории. В случае осесимметричных направленных потоков уравнения Навье-Стокса имеют следующий вид:
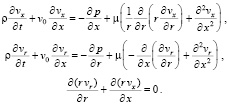
(9)
Здесь v0 – основная скорость направленного потока, вокруг которой происходит малая пульсация составляющих скоростей: vx = v0 + vʹ; vr = vʹ.
Динамические уравнения осесимметричных колебаний предварительно натянутой круглой цилиндрической оболочки записываются в виде:
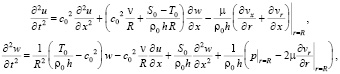
(10)
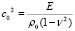
где .
Условия «прилипания» частиц жидкости к стенкам сосуда заменим условием стесненного их скольжения по поверхности контакта:
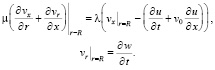
(11)
λ – коэффициент вязкого трения материала оболочки и жидкости.
Рассмотрим систему уравнений (9), (10) с контактными условиями (11). Умножим уравнение (1) на r и продифференцируем по x. Далее второе уравнение системы также умножим на r и продифференцируем по r, после чего сложим левые и правые части полученных уравнений. В результате получим уравнение для давления:
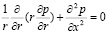
. (12)
Запишем преобразованную систему уравнений:
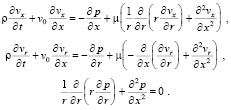
(13)
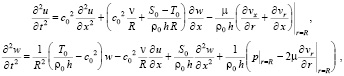
Для дальнейшего упрощения предложенной системы уравнений в трехмерной постановке можно вместо уравнений Навье-Стокса (9) использовать уравнения Эйлера для описания движения направленного потока идеальной несжимаемой жидкости, полагая при этом, что вязкое стесненное трение жидкости о стенку сосуда будет происходить в бесконечно тонком слое (погранслой) на контактной поверхности.
Читайте также: Washbalsam nordland для мембранных тканей
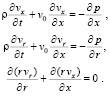
(14)
Вязкие свойства жидкости в погранслое будут описываться упрощенным одномерным уравнением, записанным на основании первого уравнения Навье-Стокса:
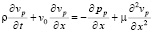
. (15)
При этом на границе погранслоя функция давления может испытывать конечный скачок. Касательные напряжения на стенке будут иметь вид:
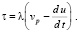
(16)
Тогда уравнения движения оболочки примут вид:
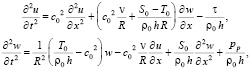
(17)
В качестве контактных условий для идеальной жидкости можно взять условия непроницаемости стенки и условия прилипания частиц жидкости вдоль стенки:
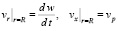
. (18)
Из системы уравнений (14)-(17) путем простых преобразований может быть получено уравнение для давления в виде (12).
Запишем преобразованную систему уравнений:
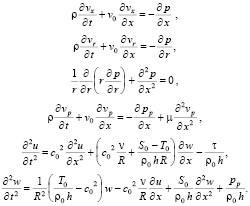
(19)
Система (19) с контактными условиями (18) представляет собой упрощенный вариант системы уравнений динамики кровотока в сосудах с упругими стенками, так как уравнения Навье-Стокса здесь заменяются уравнениями Эйлера для идеальной жидкости, которые имеют более простой вид.
Решение основной системы уравнений для случая пульсирующего кровотока как для системы (9)-(10), так и для системы (19) можно искать в виде простых гармонических волн:
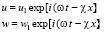
, (20)
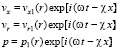
, (21)
где ω – частота пульсации кровотока, χ – волновое число.
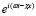
Подставляя выражения (21) в уравнения движения жидкости основной системы, получим базовые решения для амплитуд компонент скорости и давления. В этом случае сумма базовых решений для каждой волновой гармоники даст общее решение основной системы.
Неизвестные константы необходимо определять из контактных условий, уравнений движения стенки, а также из граничных условий на входе и выходах из сосудистой системы. Это напрямую связано с решением дисперсионных уравнений, возникающих при подстановке выражений (20) в основную систему.
Были получены начальные точки (рис. 1 и 2) дисперсионных кривых для дисперсионного уравнения, полученного при решении системы (1)-(7). На рисунках значения по осям а и b – это действительная и мнимая части волнового числа.
Используя полученные точки в качестве начальных приближений для построения решения дисперсионного уравнения, можно построить необходимые дисперсионные кривые, что позволит завершить решение полной краевой задачи с учетом краевых и контактных условий и найти необходимое число констант интегрирования.
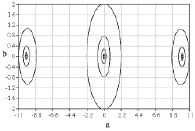
Рис. 1. Начальные точки дисперсионных кривых в случае малой вязкости
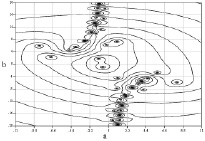
Рис. 2. Начальные точки дисперсионных кривых в случае большой вязкости
Такой алгоритм поиска общего решения системы (1)-(7) вполне подходит для решения систем (9)-(10) и (19). Таким образом, предложенные математические модели могут использоваться для исследования кровотока в системе кровеносных сосудов.
Результаты и обсуждение
Предложенные варианты постановки задач гемодинамики в системе кровеносных сосудов позволяют уменьшить количество основополагающих предположений и упростить основную систему уравнений. В случае, когда проводится учет направленных потоков жидкости, отпадает необходимость пренебрежения конвективной составляющей ускорения частиц, что повышает точность результатов.
Новый вид кинематических контактных условий стесненного скольжения (11), по мнению авторов, более адекватно описывает процесс взаимодействия потока со стенкой. В реальном сосуде в случае выполнения условий прилипания частиц неизбежным был бы процесс образования атеросклеротических отложений, который является патологическим. Таким образом, математические модели, в которых взаимодействие крови со стенкой описывается подобным образом, не могут достоверно описывать процесс гемодинамики.
Замена уравнений Навье-Стокса уравнениями Эйлера в системе (19) существенно упрощает основную систему, при этом уравнение для тонкого погранслоя вблизи стенки сосуда (15) позволяет учесть все возникающие в этой области процессы.
Таким образом, предложен ряд упрощений и уточнений, позволяющий более точно и адекватно моделировать движение крови в системе кровеносных сосудов человека.
Предложены новые варианты постановки задач о движении крови в сосудах с упругими стенками, позволяющие уменьшить количество основополагающих гипотез и, тем самым, повысить строгость и реалистичность предлагаемых математических моделей.
Кроме того, предложен новый вид кинематических контактных условий, позволяющих более точно описать процесс взаимодействия потока крови с сосудистой стенкой.
Моделирование ткани кровеносных сосудов это

Ангиогенез (а) – сложный процесс формирования новых кровеносных сосудов в органах либо тканях, предопределенный строго поочередной работой множества факторов в пространственно-временной позиции. Новые капилляры образуются из мелких кровеносных сосудов, путем активизирования эндотелиальной клетки, преобразования в них протеиназ, деградации внеклеточной структуры ткани, разрастания и передвижения клеточных структур. Впоследствии благодаря формированию клетками первичных высокопроницаемых сосудов, начинается стабилизация и «взросление» структур клетки с помощью притягивания перикапиллярных клеток и клеток гладких мышц. В результате чего строится многогранная сеть сосудов. В норме ангиогенез происходит в спокойном темпе, активируется он при условии тканевых ранений, наличия тромбов и других патологических процессах [1]. Физиологический ангиогенез – это реакция ткани на гормональную стимуляцию (ангиогенез в репродуктивной системе) или изменения в окружающей среде (в ответ на ишемию ткань может расширять сосудистую сеть). Исследования последних научных работ приходят к выводу, что главным стимулом ангиогенеза является дефицит кислорода, который вызывает гипоксию или ишемию, при этом HIF-1 содействует выразительности васкулярных факторов, а именно фактору роста внешнего сосудистого слоя VEGF и его нервных окончаний, который является основой регулирования роста сосудов в различных периодах развития организма [2]. Физиологический ангиогенез представлен реакцией адаптации к дефициту кислорода, поскольку VGEF считается стресс-индуцированным белком, регулируемый глюкозой и кислородом. Фактор роста индивидуально отбирает эндотелиальные клетки (ЭК) для активации их преобразования и передвижения. Увеличивает пропускаемость сосуда для прохода белков в периваскулярное пространство, необходимое для контролируемой миграции ЭК и развитию вазодилатации [3]. В этапе развития новой сети сосудов принимает участие проангиогенный фактор, который сдерживает эндотелиальную пролиферацию, снижает проницаемость сосуда и содействует притягиванию перикапиллярных клеток. Tie2 (тирозинкиназные рецепторы) играют ведущую роль в процессах роста, развития и дифференцировки клеток. Они вместе с ангиопоэтинами играют роль в корректировке сопряжения эндотелия с рядом лежащими клетками [4]. Для роста системы сосудов в эмбриональном периоде необходима система сигналов Tie/Ang, соединяемая с VEGF и его рецепторами, равно как и каскад сигнализации. Tie2/Ang1 является несамостоятельным, промотирующим ассоциацию перицитов и эндотелия, снижающим сосудистую проницаемость и обладающим противовоспалительной активностью каскадом сигнализации [5]. Ang1 помогает образовывать связь между перицитами и эндотелиальными клетками при связывании с экспрессируемым на поверхности клеток эндотелия рецептором Tie2, помогая стабилизации, находящегося в стадии развития сосудистой системы [6, 7]. 2. Тромбоцитарный ФР (PDGF), который привлекает перициты и ГМК. Это белок, синтезируемый в мегакариоцитах и находящийся в гранулах тромбоцита. Все элементы – это результат роста фактора около тысячи молекул тромбоцитов. Фактор – мощный стимул восстановления тканей. Рецепторы для этого находятся в стенке сосудов на поверхности фибробласта и клетках гладкой мускулатуры. PDGF активизирует пролиферацию таких клеток. Более того, PDGF усиливает выработку компонентов соединительной ткани (коллагена, гистамина и др.) [8]. 3. Трансформирующий ФР-β1 (TGF-β1) стимулирует синтез белков внеклеточной матрицы. Контролирующий пролиферацию полипептид (представитель цитокинов) в большинстве клеток также регулирует дифференциацию и другие функциональные особенности. Члены семейства TGF-β1 проявляют множественное воздействие на огромное количество видов клеток и способствуют контролю роста клеток, дифференциации и апоптоза, а также в модуляции иммунной системы [9]. Артериогенез способствует формированию коллатеральных сосудов из неактивных артериальных сетей, по которым кровь проходит в места замыкания. Главным катализатором такого процесса является увеличение напряжения сдвига выше места окклюзии, способствующего преобразованию молекул адгезии клетками эндотелия с последующей аккумуляцией моноцитов в стенке сосуда. Они секретируют функционирующие ФР, основными регуляторами артериогенеза являются фактор роста фибробластов (FGF), и PDGF, VEGF и CXC-хемокины (подсемейства, характеризующиеся наличием одной аминокислоты, которая разделяет N-концевые цистеины) [10]. Действия ангиогенеза контролируются ФР во временном пространстве, этот факт следует учесть во время терапевтического ангиогенеза. Стабильное состояние сосудистой сети в организме постнатального периода обеспечивается равноценным соотношением между активаторами ангиогенеза (в основном ФР и цитокинами) и его ингибиторами (тромбоспондином, ангиостатин, тумастин, эндостатином и др.), движение такого баланса в сторону активаторов, в большинстве случаев, непродолжительный, ведет за собой активацию ангиогенеза [11]. Примерами являются воспаление, заживление ран, ишемия.
Читайте также: Какие физические закономерности лежат в основе газообмена в тканях
Данный процесс и образование отростчатых сосудов проходит в несколько последовательных шагов. В первой фазе начинается активация перицитов, которые находятся в тесном контакте с эндотелием, они увеличиваются в объеме, укорачивая свои отростки. Таким образом, происходит ослабление межклеточных контактных соединений. Перициты проецируются в периваскулярное пространство, происходит деградация базальной мембраны и диссоциация перицитов и эндотелия [12]. Хотя процесс на начальных стадиях роста эндотелиоцитов в новообразованную васкуляризованную ткань может протекать без помощи перицитов, в последующей работе именно они локализуются по ходу прорастания эндотелия и формируют процессы, которыми направляются новообразованные сосуды [13]. Клетки эндотелиоциты берут начало своего роста в тканях по направлению Ang-1 продуцирующей ткани и, производя ферменты, катепсины и активаторы плазминогена, которые ведут к ухудшению базальной мембраны, так же матриксные металлопротеиназы (MMPs), являющиеся основными протеолитическими энзимами, принимающими участие в этом процессе. Внеклеточный матрикс, расщепляясь, формирует полипептиды, фрагменты этих белков имеют как про- так и антиангиогенные эффекты. Растворение белков внеклеточного матрикса проходит под контролем ингибиторов протеаз (UAP, PAI) [14]. При участии молекул клеточной адгезии и 17 интегринов, лигандами для которых служат белки внеклеточного матрикса (фибронектин, ламинин, витронектин), ослабление межклеточных контактных соединений эндотелиальных клеток и разрушение базальной мембраны дает начало далее следующему перемещению эндотелиальных клеток в околососудистый участок [15]. Клетки эндотелия начинают активно пролиферировать, сформировывая структуры в виде канала, далее преобразовываясь в зрелую сосудистую сеть. Некоторые сосуды микроциркуляторного русла объединяются в целостную сеть, посредством которой происходит перфузия тканей (рис. 1). До этого момента VEGF влияет на сохранение клеток эндотелия и их целостность [16].
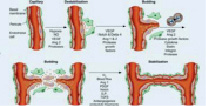
Момент подъема эндотелия сосудов. В процессе проницаемости сосудов лежит база регулировки А. VEGF – мощнейший индуктор ангиогенеза в целом ряде опытных моделей in vivo [17]. Он представляет собой гомодимерный, высокогликолизированный, митогенный белок, предназначенный для эндотелиальных клеток. Большинство ученых считают, что VEGF взаимодействует с цитокинами, которые имеют сенсоры с протеолитическими ферментами и растворимые антагонисты. Они, взаимодействуя, регулируют высвобождение цитокинов из внеклеточного матрикса [18]. Группа VEGF включает в себя ряд образцов: EGF-A способствует наращиванию проницаемости сосуда; VEGF-B регулирует спад внеклеточного матрикса, адгезии и клеточного передвижения; VEGF-C и VEGF-D играют главную роль в регуляции лимфатических кровеносных сосудов; так же VEGF-E является вирусным гомологом и способствует плацентарному подъему (PIGF). Он отвечает за построение сосудистой сети в плаценте (рис. 2).
Читайте также: Ткань для печати сумок
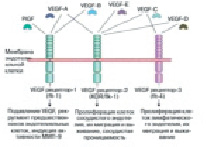
Совместное взаимодействие VEGF с рецепторами VEGFR-2 подключает активацию всех имеющихся сигнальных путей каскада. Два таких примера показаны на рисунке. Такое взаимодействие приводит к выживанию клетки и проницаемости сосудов, путем поглощения генов, которые способствуют пролиферации и передвижению клеток эндотелия. К примеру, связывание VEGF с сенсором VEGFR-2 приводит к димеризации сенсора с дальнейшей активацией пути синтеза ДНК и подъема клеток PLC-PKC-Raf-MEK-митогенактивированной белковой киназы (MAPK), а еще дальнейшей инициацией синтеза ДНК и подъема клеток, за это время как активация фосфатидилинозита 3’-киназы (PI3K)-Akt-way обязана прирастить подъем эндотелиальных клеток. Передвижение клеток и изменения в цитоскелете актина вызывает ген Src. Рецепторы VEGF располагаются на поверхности эндотелиальной клетки, но не теряют способности стать внутриклеточными [19]. Они являются участниками процесса приспособления клетки для выживания. VEGFR-2 представлен полноразмерным сенсором, прикрепляющимся к плоскости клеточки. VEGF-CcVEGFR-3 связывание опосредует лимфангиогенез. VEGF имеет возможность вязать рецепторный нейропилин (NRP), который имеет возможность работать как coreceptor с VEGFR-2 (горизонтальная стрелка 1) и имеет возможность регулировать A [20]. Комбинированное внедрение VEGF в экспериментальных исследовательских работах, а еще фактора стабилизации сосудов ангиопоэтин-1, и тромбоцитов FR (PDGF-BB) в композиции с FGF-2 вызывает возникновение сосудистой сети, которая продолжает оставаться размеренной сквозь 1 год впоследствии остановки данных моментов [21, 22]. Иным раскладом к более равновесной стимуляции ангиогенеза, вполне вероятно, может быть создание генетических структур, основанных на консистенции геномной ДНК, а еще cDNA-форм гена VEGF, имеющие в для себя экзоны и интроны в иной области слияния. Другая стратегия имеет возможность основываться на применении генов, которые кодируют моменты, которые активируют поглощение множества ангиогенных молекул [23]. Фактором, обеспечивающим больше обобщенные сигналы ангиогенеза, имеет возможность быть активатор плазминогена наподобие урокиназы (urokinase) – протеаза серина, ведущей регулятор внеклеточного протеолиза, а еще моделирование тканей. Урокиназа инициирует составление капилляров и артериол и наращивает скопление макрофагов в зоне периинфаркции, сокращаяет величину возникших, увеличивает васкуляризацию, готовит более скорым восстановление перфузии и не позволяет развиваться некрозу в ишемической конечности [24].
Дефицитность кровоснабжения приводит к гипоксии по причине понижения диффузии воздуха. Гипоксия считается более необходимым катализатором A, есть активация метаболических стезей, которые индуцируются белками, этими как момент гипоксии 1, собственно, что приводит к наращиванию экспрессии проангиогенных моментов, этих как моменты подъема VEGF и фибробластов [25]. Впоследствии подключения А случается перелом соединительнотканной пластинки и внеклеточного матрикса (ВКМ), вследствие увеличенной энергичности матрикса. Далее клетки организуются в канальцы, с просветами образуя свежую капиллярную металлопротеиназу (ММП). Во время сего процесса притягиваются перициты, которые прикрепляются к свежим кровеносным сосудам и стабилизируются. До сего этапа созревания единство и выживание эндотелиальных клеток находятся в зависимости от VEGF1.13. Другой методикой экспрессии считается втягивание воспалительных клеток, цитокинов ФНО (фактор некроза α-опухолей) и ИЛ-1, которые в собственную очередь индуцируют продукцию обычных клеток. Подъем микрососудов продолжается до тех пор, пока же не достигнется очень максимально вероятная близость к клеточке. Впоследствии А. перебегает в стадию спокойствия (в дамской репродуктивной системе ангиогенный цикл считается исключением). Каждое наращивание массы ткани сопрягается с уноваскуляризацией, которая поддерживает необходимую плотность сосудов. Например, А. индуцируется, когда метаболическая надобность выше перфузионную дееспособность имеющих место быть сосудов. По-видимому, устройство данной адаптивной реакции заключается в том, собственно, что условный недостаток воздуха приводит к ужесточению ангиогенных стимулов.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом