
Морфофункциональная характеристика мышечных тканей таблица
Это ткань энтомезенхимного происхождения, которая делится на два вида: висцеральную и сосудистую. В эмбриональном гистогенезе даже электронно-микроскопически трудно отличить мезенхимные предшественники фибробластов от гладких миоцитов. В малодифференцированных гладких миоцитах развиты гранулярная эндоплазматическая сеть, комплекс Гольджи. Тонкие филаменты ориентированы вдоль длинной оси клетки. По мере развития размеры клетки и число филаментов в цитоплазме возрастают. Постепенно объем цитоплазмы, занятый сократительными филаментами, увеличивается, расположение их становится все более упорядоченным. Пролиферативная активность гладких миоцитов в миогенезе постепенно снижается. Это происходит в результате увеличения продолжительности клеточного цикла, выхода клеток из цикла репродукции и перехода в дифференцированное состояние.
Однако и в дефинитивном состоянии в гладкой мышечной ткани клеточная регенерация в виде размножения миоцитов полностью не прекращается. Существуют данные о том, что пролиферация и дифференцировка в большей степени свойственна субпопуляции малых (по размерам) гладких миоцитов.
Строение гладкой мышечной ткани. Структура дефинитивных гладких миоцитов (лейомиоцитов), входящих в состав внутренних органов и стенки сосудов, имеет много общего, но в то же время характеризуется гетероморфией. Так, в стенках вен и артерий обнаруживаются овоидные, веретеновидные, отростчатые миоциты длиной 10-40 мкм, доходящие иногда до 140 мкм.
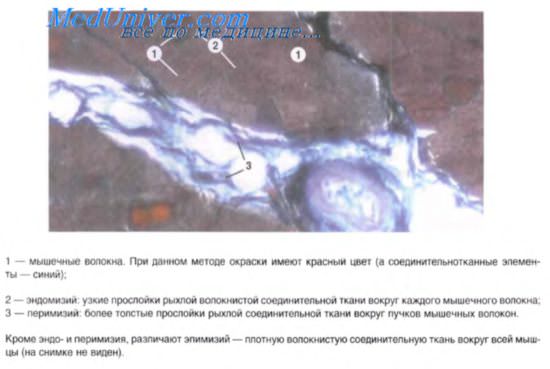
Гладкая мышечная ткань
Наибольшей длины гладкие миоциты достигают в стенке матки — до 500 мкм. Диаметр миоцитов колеблется от 2 до 20 мкм. В зависимости от характера внутриклеточных биосинтетических процессов различают контрактилъные и секреторные миоциты. Первые специализированы на функции сокращения, но вместе с тем сохраняют секреторную активность. Плазмолемма расслабленной клетки имеет ровную поверхность, а при сокращении становится складчатой. В центре клетки имеется палочковидное ядро, которое при сокращении клетки спиралевидно изгибается. Практически все ядра миоцитов содержат диплоидное количество ДНК. Гладкая эндоплазматическая сеть занимает примерно 2-7% объема цитоплазмы, а гранулярная сеть в контрактильных миоцитах выражена плохо. Митохондрии мелкие, сферические или овоидные, расположены у полюсов ядра. Характерной чертой гладких миоцитов является наличие множества впячиваний (кавеол) плазмолеммы, содержащих ионы кальция.
Секреторные миоциты (синтетические) по своей ультраструктуре напоминают фибробласты, однако содержат в цитоплазме пучки тонких миофиламентов, расположенные на периферии клетки. В цитоплазме хорошо развиты комплекс Гольджи, гранулярная эндоплазматическая сеть, много митохондрий, гранул гликогена, свободных рибосом и полисом. По степени зрелости такие клетки относят к малодифференцированным.
Сократительный аппарат миоцитов представлен тонкими актиновыми филамен-тами (гладкомышечным альфа-актином), связанными с тропомиозином. Толстые нити состоят из миозина, мономеры которого располагаются вблизи филаментов актина. Соотношение актиновых и миозиновых филаментов в гладком миоците составляет 12 к 1. Важным компонентом контрактильного аппарата миоцитов являются электронно-плотные структуры — тельца прикрепления, расположенные свободно в цитоплазме (плотные тельца) или тесно связанные с плазмолеммой. Основными белковыми компонентами плотных телец являются альфа-актинин, актин (немышечный) и кальпонин, что позволяет расссматривать их как функциональный эквивалент Z-линий миофибрилл скелетной мышцы. Актиновые филаменты фиксируются на плотных тельцах. Промежуточные филаменты, включающие десмин и виментин, обеспечивают связи между плотными тельцами и плазмолеммой, образуя прикрепительные пластины.
Сократительные белки формируют решетчатую структуру, закрепленную по окружности плазмолеммы, поэтому сокращение выражается в укорочении клетки, которая приобретает складчатую форму, тогда как в состоянии покоя клетка вытянута. При возникновении нервного импульса, распространяющегося по плазмолемме миоцита, происходит повышение уровня внутриклеточного Са2+, который поступает в цитоплазму из кавеол, отшнуровывающихся в цитоплазму в виде пузырьков. Высвобождение ионов кальция приводит к каскаду реакций, в результате которого происходит полимеризация миозина и образование перекрестных связей миозина вдоль актиновых филаментов по мере развития мышечного сокращения. Расслабление мышцы возникает при восстановлении концентрации исходного уровня Са2+ внутри клетки путем его перемещения внутрь саркоплазматической сети. При этом образовавшиеся в присутствии ионов кальция связи между актином и миозином нарушаются, акто-миозиновый комплекс распадается, гладкий миоцит расслабляется.
Читайте также: Ткани бифлекс в с петербурге
Гладкие миоциты синтезируют протеогликаны, гликопротеиды, проколлаген, проэластин, из которых формируются коллагеновые и эластические волокна и основное вещество межклеточного матрикса.
Взаимодействие миоцитов осуществляется с помощью цитоплазматических мостиков, взаимных впячиваний, нексусов, десмосом или простых участков мембранных контактов клеточных поверхностей.
Регенерация гладкой мышечной ткани
Гладкая мышечная ткань висцерального и сосудистого видов обладает значительной чувствительностью к воздействию экстремальных факторов.
В активированных миоцитах возрастает уровень биосинтетических процессов, морфологическим выражением которых являются синтез сократительных белков, укрупнение и гиперхроматоз ядра, гипертрофия ядрышка, возрастание показателей ядерно-цитоплазменного отношения, увеличение количества свободных рибосом и полисом, активация ферментов, аэробного и анаэробного фосфорилирования, мембранного транспорта. Клеточная регенерация осуществляется как за счет дифференцированных клеток, обладающих способностью вступать в митотический цикл, так и за счет активизации камбиальных элементов (миоцитов малого объема).
При действии ряда повреждающих факторов отмечается фенотипическая трансформация контрактильных миоцитов в секреторные. Данная трансформация часто наблюдается при повреждении интимы сосудов, формировании ее гиперплазии при развитии атеросклероза.
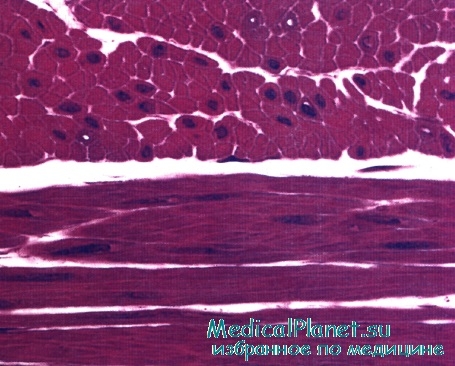
Гладкая мышечная ткань в поперечном (наверху) и продольном (внизу) разрезах. Обратите внимание на центрально расположенные ядра. Во многих клетках ядра не попали в срез.
Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
1. Мышечные ткани. Источники развития. Общая морфо-функциональная характеристика. Классификация. Возможности регенерации.
Мышечными тканями называют ткани, различные по строению и происхождению, но сходные по способности к выраженным сокращениям. Они обеспечивают перемещения в пространстве организма в целом, его частей и движение органов внутри организма.
Основные морфологические признаки элементов мышечных тканей- удлиненная форма, наличие продольно расположенных миофибрилл и миофиламентов- специальных органелл, обеспечивающих сократимость, расположение митохондрий рядом с сократительными элементами, наличие включений гликогена, липидов и миоглобина.
Специальные сократительные органеллы- миофиламенты или миофибриллы обеспечивают сокращение, которое возникает при взаимодействии в них двух основных фибриллярных белков- актина и миозина при обязательном участии ионов кальция . Митохондрии обеспечивают эти процессы энергией. Запас источников энергии образует гликоген липиды. Миоглобин- белок, обеспечивающий связывание кислорода и создание его запаса на момент сокращения мышцы, когда сдавливаются кровеносные сосуда.
Классификация.В соответствии с морфофункциональным принципом, в зависимости от структуры органелл сокращения, мышечные ткани подразделяют на две подгруппы.
Первая подгруппа- поперечнополосатые мышечные ткани.
Вторая подгруппа – гладкие мышечные ткани.
В соответствии с гистогенетическим принципом в зависимости от источников развития мышечные ткани подразделяются на 5 типов: мезенхимные( из десмального зачатка в составе мезенхимы), эпидермальные (из кожной эктодермы и из прехордальной пластинки), нейральные ( из нервной трубки), целомические( из миоэпикардиальной пластинки висцерального листка сомита) и соматические(миотомные).
Первые три типа относятся к подгруппе гладких мышечных тканей, четвертый и пятый- к подгруппе поперечнополосатых.
2. Гладкая мышечная ткань. Источник развития. Морфо- функциональная характеристика гладких мышечных тканей. Структурные основы сокращения. Иннервация. Регенерация.
Различают три группы гладких мышечных тканей- мезенхимные, эпидермальные и нейральные.
Стволовые клетки и клетки – предшественники в гладкой мышечной ткани на этапах эмбрионального развития пока точно не отождествлены. Они мигрируют к местам закладки органов, будучи уже детерминированными. Дифференцируясь, они синтезируют компоненты матрикса и коллагена базальной мембраны, а также эластина. У дефинитивных клеток синтетическая способности снижена, но не исчезает полностью.
Читайте также: Зеленый алтай средство для стирки одежды из мембранных тканей
Гладкий миоцит- веретеновидная клетка длиной 20-500 мкм, шириной 5-8 мкм. Ядро палочковидное, находится в ее центральной части. Когда миоцит сокращается, его ядро изгибается и даже закручивается. Филаменты актина образуют в цитоплазме трехмерную сеть, вытянутую преимущественно продольно. Концы филаментов скреплены между собой и с плазмолеммой специальными сшивающими белками. Мономеры миозина располагаются рядом с филаментами актина. Сигнал к сокращению обычно поступает по нервным волокнам. Медиатор, который выделяется из их терминалей, изменяет состояние плазмолеммы. Она образует выпячивания- кавеолы,в которых концентрируются ионы кальция. Кавеолы отшнуровываются в сторону цитоплазмы в виде пузырьков. Это влечет за собой взаимодействие миозина с актином. Актиновые филаменты смещаются друг другу навстречу, плотные пятна сближаются, усилие передается на плазмолемму, и вся клетка укорачивается. Когда поступление сигналов со стороны нервной системы прекращается, ионы кальция эвакуируются из кавеол, миозин деполимеризуется и « миофибриллы» распадаются. Сокращение прекращается. Гладкая МТ иннервируется вегетативной нервной системой, т.е. не подчиняется воле человека. Сокращение ГМТ медленное — тоническое, зато ГМТ малоутомляема. ГМТ в эмбриональном периоде развивается из мезенхимы. Вначале мезенхимные клетки имеют звездчатую, отросчатую форму, а при дифференцировке в ГМ-клетки приобретают веретеновидную форму; в цитоплазме накапливаются органоиды спецназначения — миофибриллы из актина и миозина. Регенерация ГМТ: 1. Митоз миоцитов после дедифференцировки: миоциты утрачивают сократительные белки, исчезают митохондрии и превращаются в миобласты. Миобласты начинают размножаться, а потом вновь дифференцируются в зрелые леомиоциты. 2. Возможно образование новых ГМ-клеток из малодифференцированных стволовых клеток фибробластического дифферона рыхлой с.д.Физиологическая регенерация гладкой мышечной ткани проявляется в условиях повышенных функциональных нагрузок. Наиболее отчетливо это видно в мышечной оболочке матки при беременности. Такая регенерация осуществляется не столько на тканевом, сколько на клеточном уровне: миоциты растут, в цитоплазме активизируются синтетические процессы, количество миофиламентов увеличивается. Не исключена, однако, и пролиферация клеток.
3. Скелетная поперечно-полосатая мышечная ткань. Источники развития. Морфо-функциональная характеристика миосимпласта. Структурные основы сокращения. Мышца как орган. Типы мышечных волокон. Регенерация.
Источником развития элементов скелетной поперечнополосатой мышечной ткани являются клетки миотомов- миобласты.Одни из них дифференцируются на месте и участвуют в образовании так называемых аутохтонных мышц. Другие клетки мигрируют из миотомов в мезенхиму. В ходе дифференцировки возникают две клеточные линии. Клетки одной из линий сливаются, образуя удлиненные сиппласты-мышечные трубочки.
В них происходит дифференцировка специальных органелл- миофибрилл. Клетки другой линии остаются самостоятельными и дифференцируются в миосимпластов. Основной структурной единицей скелетной мышечной ткани является мышечное волокно, состоящие из миосимпласта миосателлитоцитов, покрытых общей базальной мембраной.
Миосимпласт имеет множество продолговатых ядер, расположенных непосредственно под сарколеммой. Их количество в одном симпласте может достигать нескольких десятков тысяч. Миофибриллы заполняют основную часть миосимпласта и расположены продольно.
Типы мышечных волокон. Мышечные волокна в составе разных мышц обладают разной силой, скоростью и длительностью сокращения, а также утомляемостью. Ферменты в них обладают разной активностью и представлены в различных изомерных формах. Заметно различие в них содержания дыхательных ферментов- гликолитических и окислительных. По соотношению миофибрилл, митохондрий и миоглобина различаютбелые, красные и промежуточные волокна. По функциональным особенностям мышечные волокна подразделяют набыстрые, медленные, и промежуточные.Обычно в быстрых волокнах преобладают гликолитические процессы, они более богатыгликогеном, в них меньше миоглобина, поютому их называют такжебелыми.В медленных волокнах, напротив, выше активность окислительных ферментов, они богачемиоглобином, выглядят более красными.
Читайте также: Соэ при саркоме мягких тканей
Регенерация. Ядра миосимпластов делиться не могут, так как у них отсутствуют клеточные центры. Камбиальными элементами служат миосателлитоциты. Пока организм растет, они делятся, а дочерние клетки встраиваются в концы симпластов. После повреждения мышечного волокна на некотором протяжении от места травмы оно разрушается и его фрагменты фагоцитируются макрофагами. Восстановление тканей осуществляется за счет двух механизмов: компенсаторной гипертрофии самого симпласта и пролифирации миосателлитоцитов. В симпласте активизируются гранулярная эндоплазматическая сеть и аппарат Гольджи. Поврежденный конец миосимпластаутолщается, образуя мышечную почку.Миосателлитоциты, сохранившиеся рядом с повреждением, делятся. Одни из них мигрируют к мышечной почке и встраиваются в нее, другие сливаются и образуют миотубы, которые затем входят в состав вновь образованных мышечных волокон или формируют новые волокна.
Скелетная мышца как орган.Передача усилий сокращения на скелет осуществляется посредством сухожилий или прикрепления мышц непосредственно к надкостнице.На конце каждого мышечного волокна плазмолемма образует глубокие узкие впячивания.В них со стороны сухожилия или надкостницы проникают тонкиеколлагеновые волокна. Последние спирально оплетаются ретикулярными волокнами. Концы волокон направляются к базальной мембране, входят в неё, поворачивают назад и по выходе снова оплетают коллагеновые волокна соед.ткани.Между мышечными волокнами находятся тонкие прослойки рыхлой волокнистой соед.ткани— эндомизий.Более толстые прослойки рыхлой соединт.ткани окружают по несколько мышечных волокон,образуяперемизийи разделяя мышцу на пучки. Соединительную ткань ,окружающую поверхность мышцы, называютэпимизием.
4. Сердечная поперечно-полосатая мышечная ткань. Источник развития. Морфофункциональная характеристика кардиомиоцита. Структурные основы совращения. Морфофункциональная характеристика поперечно-полосатой сердечной мышечной ткани.
Гистогенез и виды клеток. Источники развития поперечно-полосатой мышечной ткани-симметричные участки висцерального листка спланхнотома в шейной части зародыша—миоэпикардиальные пластинки.Из них дифференцируются также клетки мезотелия эпикарда. В ходе гистогенеза возникает 5 видов кардиомиоцитов- рабочие, синусные переходные, проводящие, а также секреторные.
Рабочие кардиомиоциты образуют свои цепочки. Именно они, укорачиваясь, обеспечивают силу сокращения всей сердечной мышцы. Рабочие кардиомиоциты способны передавать управляющие сигналы друг другу. Синусные кардиомиоциты способны автоматически в определенном ритме сменять состояние сокращения на состояние расслабления. Синусныекардиомиоциты передают управляющие сигналыпереходнымкардиомиоцитам, а последние-проводящим.Проводящие кардиомиоциты образуют цепочки клеток, соединенных своими концами. Первая клетка в цепочке воспринимает управляющие сигналы от синусных кардиомиоцитов и передает их далее- другим проводящим кардиомиоцитов. Клетки, замыкающие цепочку, передают сигнал через переходные кардиомиоциты рабочим. Секреторные кардиомиоциты вырабатываютнатрийуретический фактор, участвующий в процессах регуляции мочеобразования и в некоторых других процессах. Все кардиомиоциты покрыты базальной мембраной.
Строение сократительных ( рабочих) кардиомиоцитов. Клетки имеют удлиненную форму, близкую к цилиндрической Их концы соединяются друг с другом, так что цепочки клеток составляют так называемые функциональные волокна. В области контактов клеток образуются так называемые вставочные диски. Ядро кардиомиоцита овальное и лежит в центральной части клетки. У полюсов ядра сосредоточены немногочисленные органеллы общего значения. Специальные органеллы, которые обеспечивают сокращение, называются миофибриллами.Кардиомиоциты соединяются друг с другом своими торцевыми концами образуются вставочные диски. Поперечные участки выступов соседних клеток соединены друг с другом интердигитациями и десмосомами. К каждой десмосоме со стороны цитоплазмы подходит миофибрилла, закрепляющаяся концом в десмоплакиновом комплексе. Таким образом, при сокращении тяга одного кардиомиоцита передается другому. Боковые поверхности выступов кардиомиоцитов объединяются нексусами. Это создает между ними метаболические связи и обеспечивает синхронность сокращений.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом