
Выделяют несколько последовательных этапов запуска и осуществления мышечного сокращения.
1. Потенциал действия распространяется вдоль двигательного нервного волокна до его окончаний на мышечных волокнах.
2. Каждое нервное окончание секретирует небольшое количество нейромедиатора ацетилхолина.
3. Ацетилхолин действует на ограниченную область мембраны мышечного волокна, открывая многочисленные управляемые ацетилхолином каналы, проходящие сквозь белковые молекулы, встроенные в мембрану.
4. Открытие управляемых ацетилхолином каналов позволяет большому количеству ионов натрия диффундировать внутрь мышечного волокна, что ведет к возникновению на мембране потенциала действия.
5. Потенциал действия проводится вдоль мембраны мышечного волокна так же, как и по мембране нервного волокна.
6. Потенциал действия деполяризует мышечную мембрану, и большая часть возникающего при этом электричества течет через центр мышечного волокна. Это ведет к выделению из саркоплазматического ретикулума большого количества ионов кальция, которые в нем хранятся.
7. Ионы кальция инициируют силы сцепления между актиновыми и миозиновыми нитями, вызывающие скольжение их относительно друг друга, что и составляет основу процесса сокращения мыщц.
8. Спустя долю секунды с помощью кальциевого насоса в мембране саркоплазматического ретикулума ионы кальция закачиваются обратно и сохраняются в ретикулуме до прихода нового потенциала действия. Удаление ионов кальция от миофибрилл ведет к прекращению мышечного сокращения.
Далее мы обсудим молекулярные механизмы этого процесса.
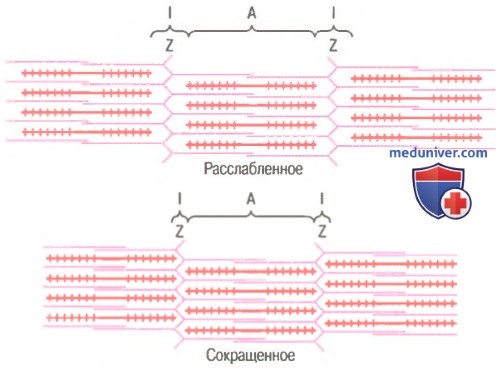
Миофибрилла в расслабленном и сокращенном состоянии. Показано (вверху), что актиновые нити (розовые) вдвинуты в пространства между миозиновыми нитями (красные). Сближение Z-дисков друг с другом (внизу).
Молекулярные механизмы мышечного сокращения
Механизм скольжения нитей для мышечного сокращения. На рисунке показан основной механизм мышечного сокращения. Показано расслабленное состояние саркомера (вверху) и сокращенное состояние (внизу). В расслабленном состоянии концы актиновых нитей, отходящие от двух последовательных Z-дисков, лишь незначительно перекрываются. Наоборот, в сокращенном состоянии актиновые нити втягиваются внутрь между миозиновыми так сильно, что их концы максимально перекрывают друг друга. При этом Z-диски притягиваются актиновыми нитями к концам миозиновых. Таким образом, мышечное сокращение осуществляется путем механизма скольжения нитей.
Что заставляет нити актина скользить внутрь среди нитей миозина? Это связано с действием сил, генерируемых при взаимодействии поперечных мостиков, исходящих от нитей миозина, с нитями актина. В условиях покоя эти силы не проявляются, однако распространение потенциала действия вдоль мышечного волокна приводит к выделению из саркоплазматическо-го ретикулума большого количества ионов кальция, которые быстро окружают миофи-бриллы. В свою очередь, ионы кальция активируют силы взаимодействия между нитями актина и миозина, в результате начинается сокращение. Для осуществления процесса сокращения необходима энергия. Ее источником являются высокоэнергетические связи молекулы АТФ, которая разрушается до АДФ с высвобождением энергии. В следующих разделах мы приведем известные детали молекулярных процессов сокращения.
Молекулярные особенности сократительных нитей
Миозиновая нить. Она состоит из множества молекул миозина, молекулярная масса каждой составляет около 480000. На рисунке показана отдельная молекула; и также — объединение многих молекул миозина в миозиновую нить, а также взаимодействие одной стороны этой нити с концами двух актиновых нитей.
В состав молекулы миозина входят 6 полипептидных цепей: 2 тяжелые цепи с молекулярной массой около 200000 каждая и 4 легкие цепи с молекулярной массой около 20000 каждая. Две тяжелые цепи спирально закручиваются вокруг друг друга, формируя двойную спираль, которую называют миозиновым хвостом. С одного конца обе цепи изгибаются в противоположных направлениях, формируя глобулярную полипептидную структуру, называемую миозиновой головкой. Таким образом, на одном конце двойной спирали молекулы миозина образуются 2 свободные головки; 4 легкие цепи также включены в состав миозиновой головки (по 2 в каждой). Они помогают регулировать функцию головки во время мышечного сокращения.
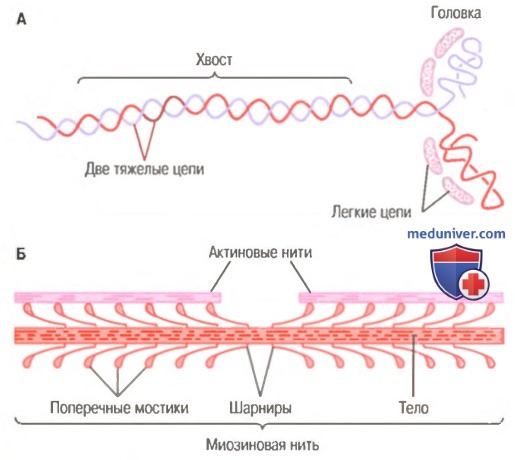
А. Молекула миозина.
Б. Объединение многих молекул миозина в одну миозиновую нить.
Показаны также тысячи миозиновых поперечных мостиков и взаимодействие их головок с прилежащими актиновыми нитями.
Миозиновая нить состоит из 200 или более отдельных молекул миозина. Видно, что хвосты молекул миозина объединяются, формируя тело нити, а многочисленные головки молекул выдаются наружу по сторонам тела. Кроме того, наряду с головкой в сторону выступает часть хвоста каждой миозиновой молекулы, образуя плечОу которое выдвигает головку наружу от тела, как показано на рисунке. Выступающие плечи и головки вместе называют поперечными мостиками. Каждый поперечный мостик может сгибаться в двух точках, называемых шарнирами. Один из них расположен в месте, где плечо отходит от тела миозиновой нити, а другой — где головка крепится к плечу. Движение плеча позволяет головке или выдвигаться далеко наружу от тела миозиновой нити, или приближаться к телу. В свою очередь, повороты головки участвуют в процессе сокращения, что обсуждается в следующих разделах.
Читайте также: Ткани для струящихся юбок
Общая длина каждой миозиновой нити остается постоянной и равна почти 1,6 мкм. В самом центре миозиновой нити на протяжении 0,2 мкм поперечных мостиков нет, поскольку снабженные шарнирами плечи отходят в стороны от центра.
Сама миозиновая нить сплетена таким образом, что каждая последующая пара поперечных мостиков смещена в продольном направлении относительно предыдущей на 120°, что обеспечивает распределение поперечных мостиков во всех направлениях вокруг нити.
АТФ-азная активность миозиновой головки. Есть и другая особенность миозиновой головки, необходимая для мышечного сокращения: миозиновая головка функционирует как фермент АТФ-аза. Как объясняется далее, это свойство позволяет головке расщеплять АТФ и использовать энергию расщепления высокоэнергетической связи для процесса сокращения.
Актиновая нить. Актиновая нить состоит из трех белковых компонентов: актина, тропомиозина и тропонина.
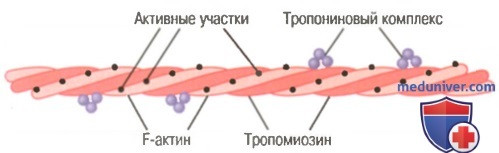
Актиновая нить, состоящая из двух спиралевидных цепочек молекул F-актина и двух цепочек молекул тропомиозина, расположенных в желобках между цепочками актина.
К одному концу каждой молекулы тропомиозина прикреплен тропониновый комплекс, который запускает сокращение.
Основой актиновой нити являются две цепи белковой молекулы F-актина. Обе цепи закручиваются в спираль так же, как и молекула миозина.
Каждая цепь двойной спирали F-актина состоит из полимеризованных молекул G-актина с молекулярной массой около 42000. К каждой молекуле G-актина прикреплена 1 молекула АДФ. Полагают, что эти молекулы АДФ являются активными участками на актиновых нитях, с которыми взаимодействуют поперечные мостики миозиновых нитей, обеспечивая мышечное сокращение. Активные участки на обеих цепях F-актина двойной спирали расположены со смещением таким образом, что вдоль всей поверхности актиновой нити встречается один активный участок примерно через каждые 2,7 нм.
Длина каждой актиновой нити — около 1 мкм. Основания актиновых нитей прочно встроены в Z-диски; концы этих нитей выступают в обоих направлениях, располагаясь в пространствах между миозиновыми молекулами.
Молекулы тропомиозина. Актиновая нить также содержит другой белок — тропомиозин. Каждая молекула тропомиозина имеет молекулярную массу 70000 и длину 40 нм. Эти молекулы спирально оплетают спираль из F-актина. В состоянии покоя молекулы тропомиозина располагаются поверх активных участков актиновых нитей, препятствуя их взаимодействию с миозиновыми нитями, лежащему в основе сокращения.
Тропонин и его роль в мышечном сокращении. По ходу молекул тропомиозина к ним периодически прикреплены другие белковые молекулы, называемые тропонином. Они представляют собой комплексы трех слабосвязанных белковых субъединиц, каждая из которых играет специфическую роль в регуляции мышечного сокращения. Одна из субъединиц (тропонин I) имеет высокое сродство к актину, другая (тропонин Т) — к тропомиозину, третья (тропонин С) — к ионам кальция. Считают, что этот комплекс прикрепляет тропомиозин к актину. Высокое сродство тропонина к ионам кальция, как полагают, инициирует процесс сокращения, о чем говорится в следующей статье.
Видео физиология мышц и мышечного сокращения — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в оглавление раздела «Физиология человека.»
Мышечная ткань характеристика процесса сокращения
Гладкие мышцы содержат актиновые и миозиновые нити, имеющие химические характеристики, подобные актиновым и миозиновым нитям скелетных мышц. Но в гладких мышцах нет тропонинового комплекса, необходимого для запуска сокращения скелетной мышцы, следовательно, механизм инициации сокращения в них другой. Этот механизм подробно обсуждается далее в нашей статье.
Химические исследования показали, что актиновые и миозиновые нити, извлеченные из гладких мышц, взаимодействуют друг с другом во многом так же, как и в скелетной мышце. Более того, процесс сокращения активируется ионами кальция, а энергия для сокращения обеспечивается разрушением АТФ до АДФ.
Существуют, однако, значительные различия в морфологической организации гладких и скелетных мышц, а также в сопряжении возбуждения и сокращения, механизме запуска ионами кальция сократительного процесса, длительности сокращения и количестве энергии, необходимой для сокращения.
Читайте также: Искусственный снег из ткани
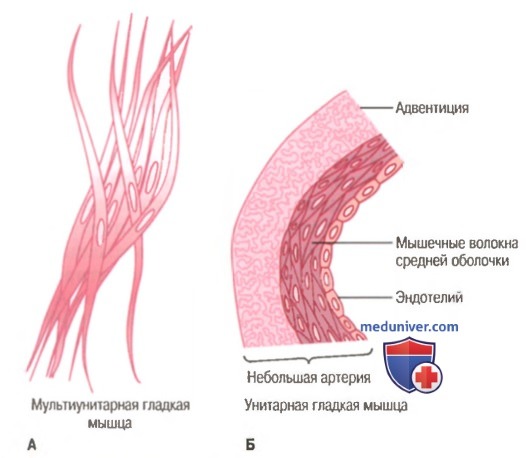
Мультиунитарная (А) и унитарная (Б) гладкие мышцы.
Морфологическая основа сокращения гладких мышц
Гладкие мышцы не имеют такой упорядоченной организации актиновых и миозиновых нитей, которая обнаруживается в скелетных мышцах, придавая им «полосатость». С помощью техники электронной микрофотографии выявляется гистологическая организация. Видно большое число актиновых нитей, прикрепленных к так называемым плотным тельцам. Некоторые из этих телец прикрепляются к клеточной мембране, другие распределяются внутри клетки. Некоторые из мембранных плотных телец соседних клеток связываются вместе мостиками из внутриклеточных белков. Через эти мостики в основном передается сила сокращения от одной клетки к другой.
В мышечном волокне среди актиновых нитей разбросаны миозиновые нити. Их диаметр более чем в 2 раза превышает диаметр актиновых нитей. На электронных микрофотографиях актиновых нитей обычно обнаруживают в 5-10 раз больше, чем миозиновых.
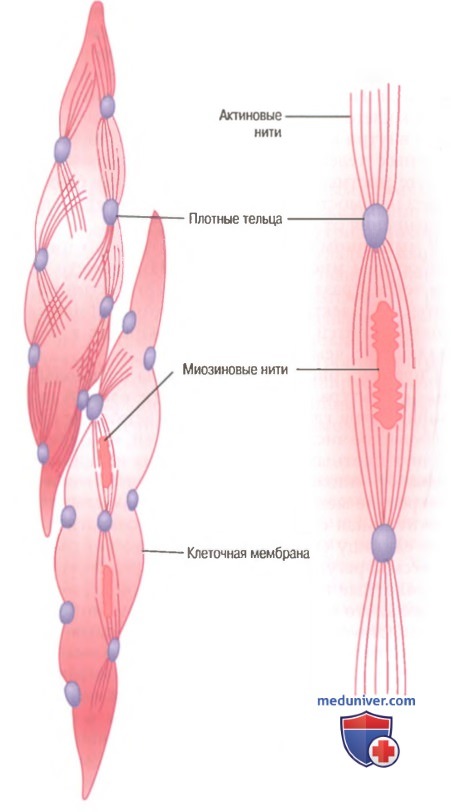
Морфологическая структура гладкой мышцы. В волокне слева вверху видны актиновые нити, исходящие из плотных телец. В волокнах слева внизу и справа на рисунке показано взаимоотношение между миозиновыми и актиновыми нитями.
На рисунке представлена предполагаемая структура отдельной сократительной единицы внутри гладкомышечной клетки, где видно большое число актиновых нитей, исходящих от двух плотных телец; концы этих нитей перекрывают миозиновую нить, расположенную посередине между плотными тельцами. Эта сократительная единица похожа на сократительную единицу скелетной мышцы, но без специфической регулярности ее структуры. В сущности, плотные тельца гладкой мышцы играют ту же роль, что и Z-диски в скелетной мышце.
Существует и другое различие. Большинство миозиновых нитей имеют поперечные мостики с так называемой боковой полярностью. Мостики организованы следующим образом: на одной стороне они шарнирно фиксируются в одном направлении, а на другой — в противоположном направлении. Это позволяет миозину тянуть актиновую нить с одной стороны в одном направлении, одновременно продвигая с другой стороны другую актиновую нить в противоположном направлении. Такая организация позволяет гладкомышечным клеткам сокращаться с укорочением до 80% их длины вместо укорочения менее чем на 30%, характерного для скелетной мышцы.
Большинство скелетных мышц сокращаются и расслабляются быстро, но сокращения гладких мышц в основном являются длительными тоническими сокращениями, которые иногда продолжаются в течение нескольких часов или даже дней. Следовательно, можно ожидать, что морфологические и химические особенности гладких мышц должны отличаться от соответствующих характеристик скелетных мышц. Далее обсуждаются некоторые из этих отличий.
Медленная циклическая активность миозиновых поперечных мостиков. В гладкой мышце по сравнению соскелетной гораздо меньше скорость циклической активности миозиновых поперечных мостиков, т.е. скорость их прикрепления к актину, отсоединение от актина и повторное прикрепление для осуществления следующего цикла. Фактически частота циклов составляет лишь от 1/10 до 1/300 этого показателя в скелетной мышце. Однако, как считают, в гладкой мышце значительно больше относительное количество времени, в течение которого поперечные мостики остаются прикрепленными к актиновым нитям, что является главным фактором, определяющим силу сокращения. Возможной причиной медленного циклирования является гораздо меньшая по сравнению со скелетной мышцей АТФ-азная активность головок поперечных мостиков, в связи с чем скорость разрушения АТФ — источника энергии для движения головок поперечных мостиков — значительно снижена с соответствующим замедлением скорости их циклов.
Видео физиология мышц и мышечного сокращения — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Мышечная ткань характеристика процесса сокращения
Эффективность двигателя или автомашины рассчитывают как процент потребляемой энергии, которая превращается в работу вместо тепла. В мышцах количество энергии, способной превращаться в работу, даже при наилучших условиях составляет менее 25% всей энергии, доставляемой к мышце (химической энергии питательных веществ), а остальная энергия превращается в тепло. Причина этой низкой эффективности связана с тем, что примерно половина энергии питательных веществ теряется во время образования АТФ, и только 40-45% энергии самой АТФ может позднее превратиться в работу.
Максимальная эффективность реализуется лишь при условии сокращения мышцы с умеренной скоростью. При медленном сокращении мышцы или без какого-либо ее укорочения во время сокращения освобождается небольшое количество поддерживающего тепла, хотя работа практически не выполняется, что снижает эффективность преобразования до нуля. Напротив, если сокращение слишком быстрое, большая доля энергии используется на преодоление вязкого трения внутри самой мышцы, и это также снижает эффективность сокращения. Обычно максимальная эффективность развивается, когда скорость сокращения составляет около 30%.
Читайте также: Джоггеры из мягкой ткани мужской
Характеристики сокращения целой мышцы
Многие особенности сокращения мышцы можно продемонстрировать на примере одиночных мышечных сокращений. Такие сокращения вызывают с помощью одиночного электрического возбуждения, иннервирующего мышцу нерва, или короткого электрического раздражения самой мышцы, что ведет к развитию одиночного сокращения, продолжающегося долю секунды.
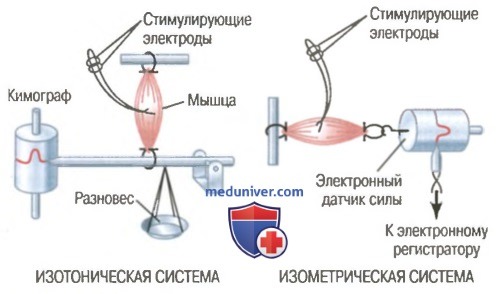 Изотоническая и изометрическая системы для регистрации мышечного сокращения.
Изотоническая и изометрическая системы для регистрации мышечного сокращения. 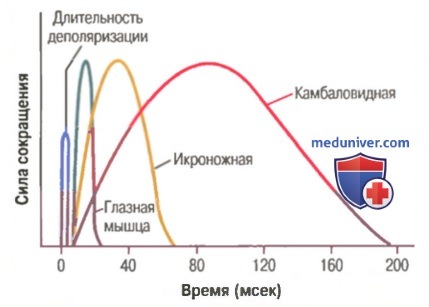 Длительность изометрических сокращений различных типов скелетных мышц млекопитающих. Показан также латентный период между потенциалом действия (деполяризацией) и мышечным сокращением.
Длительность изометрических сокращений различных типов скелетных мышц млекопитающих. Показан также латентный период между потенциалом действия (деполяризацией) и мышечным сокращением.
Изометрическое и изотоническое сокращение. Мышечное сокращение называют изометрическим, если мышца не укорачивается во время сокращения, и изотоническим — если мышца укорачивается, но ее напряжение на протяжении всего сокращения остается постоянным.
В изометрической системе мышца сокращается без уменьшения своей длины, а в изотонической системе мышца укорачивается против фиксированной нагрузки: мышца поднимает чашу весов с разновесом. Изометрическая система строго регистрирует изменения силы самого мышечного сокращения, а параметры изотонического сокращения зависят от нагрузки, против которой мышца сокращается, а также от инерции нагрузки. В связи с этим при сравнении функциональных особенностей различных типов мышц чаще всего используют изометрическую систему.
Особенности одиночных изометрических сокращений, зарегистрированных от разных мышц. В теле человека имеются много мышц разного размера — от очень маленькой стременной мышцы в среднем ухе, длиной в несколько миллиметров и диаметром около 1 мм, до очень большой четырехглавой мышцы, в 500000 раз крупнее стременной. При этом диаметр волокон может быть маленьким (10 мкм) или большим (80 мкм). Наконец, энергетика мышечных сокращений значительно варьирует от одной мышцы к другой. Поэтому не удивительно, что механические характеристики сокращений разных мышц различаются.
На рисунке показаны кривые регистрации изометрических сокращений трех типов скелетных мышц: глазной мышцы (длительность изометрического сокращения менее 1/40 сек), икроножной мышцы (длительность сокращения около 1/15 сек) и камбаловиднй мышцы (длительность сокращения примерно 1/3 сек). Интересно, что эти длительности сокращений приспособлены к функциям соответствующих мышц. Движения глаз должны быть чрезвычайно быстрыми, чтобы поддерживать фиксацию глаз на объекте для обеспечения ясного видения. Икроножная мышца должна сокращаться умеренно быстро, чтобы обеспечить скорость движения нижней конечности, достаточную для бега или прыжков. А камбаловидная мышца имеет дело в основном с медленными сокращениями для непрерывной длительной поддержки тела против силы тяжести.
Быстрые и медленные мышечные волокна. Как обсуждается в предыдущих статьях, посвященных спортивной физиологии, каждая мышца тела состоит из совокупности так называемых быстрых и медленных мышечных волокон, а также других волокон с переходными свойствами. В состав быстрореагирующих мышц входят в основном быстрые волокна и лишь небольшое число медленных. И наоборот, медленнореагирующие мышцы составлены главным образом из медленных волокон. Различия между этими двумя типами волокон следующие.
Быстрые волокна: (1) крупные волокна, обеспечивающие большую силу сокращения; (2) имеют хорошо развитый саркоплазматический ретикулум для быстрого выделения ионов кальция, инициирующих сокращение; (3) содержат большое количество гликолитических ферментов для быстрого освобождения энергии путем гликолиза; (4) имеют сравнительно бедное кровоснабжение, поскольку окислительный метаболизм имеет второстепенное значение; (5) содержат немного митохондрий также в связи со второстепенностью окислительного метаболизма.
Медленные волокна: (1) более мелкие волокна; (2) иннервируются также более мелкими нервными волокнами; (3) имеют хорошо развитую систему кровеносных сосудов и капилляров для доставки большого количества кислорода; (4) содержат значительно больше митохондрий для обеспечения высоких уровней окислительного метаболизма; (5) содержат большое количество миоглобина — железосодержащего белка, подобного гемоглобину эритроцитов. Миоглобин связывается с кислородом и хранит его до момента, когда в нем возникнет потребность (это также значительно увеличивает скорость транспорта кислорода в митохондрии). Миоглобин придает медленным волокнам красноватый вид, поэтому их называют красными волокнами, а из-за дефицита красного миоглобина в быстрых волокнах их называют белыми волокнами.
Видео физиология мышц и мышечного сокращения — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021