
Это ткань энтомезенхимного происхождения, которая делится на два вида: висцеральную и сосудистую. В эмбриональном гистогенезе даже электронно-микроскопически трудно отличить мезенхимные предшественники фибробластов от гладких миоцитов. В малодифференцированных гладких миоцитах развиты гранулярная эндоплазматическая сеть, комплекс Гольджи. Тонкие филаменты ориентированы вдоль длинной оси клетки. По мере развития размеры клетки и число филаментов в цитоплазме возрастают. Постепенно объем цитоплазмы, занятый сократительными филаментами, увеличивается, расположение их становится все более упорядоченным. Пролиферативная активность гладких миоцитов в миогенезе постепенно снижается. Это происходит в результате увеличения продолжительности клеточного цикла, выхода клеток из цикла репродукции и перехода в дифференцированное состояние.
Однако и в дефинитивном состоянии в гладкой мышечной ткани клеточная регенерация в виде размножения миоцитов полностью не прекращается. Существуют данные о том, что пролиферация и дифференцировка в большей степени свойственна субпопуляции малых (по размерам) гладких миоцитов.
Строение гладкой мышечной ткани. Структура дефинитивных гладких миоцитов (лейомиоцитов), входящих в состав внутренних органов и стенки сосудов, имеет много общего, но в то же время характеризуется гетероморфией. Так, в стенках вен и артерий обнаруживаются овоидные, веретеновидные, отростчатые миоциты длиной 10-40 мкм, доходящие иногда до 140 мкм.
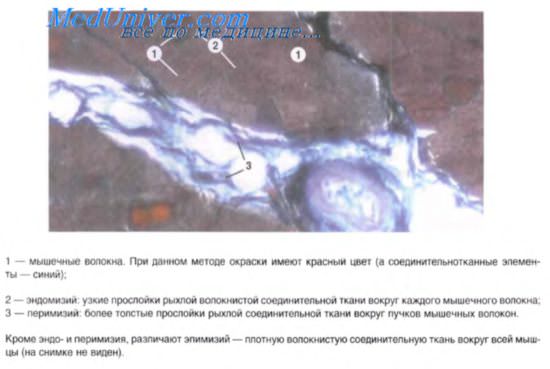
Гладкая мышечная ткань
Наибольшей длины гладкие миоциты достигают в стенке матки — до 500 мкм. Диаметр миоцитов колеблется от 2 до 20 мкм. В зависимости от характера внутриклеточных биосинтетических процессов различают контрактилъные и секреторные миоциты. Первые специализированы на функции сокращения, но вместе с тем сохраняют секреторную активность. Плазмолемма расслабленной клетки имеет ровную поверхность, а при сокращении становится складчатой. В центре клетки имеется палочковидное ядро, которое при сокращении клетки спиралевидно изгибается. Практически все ядра миоцитов содержат диплоидное количество ДНК. Гладкая эндоплазматическая сеть занимает примерно 2-7% объема цитоплазмы, а гранулярная сеть в контрактильных миоцитах выражена плохо. Митохондрии мелкие, сферические или овоидные, расположены у полюсов ядра. Характерной чертой гладких миоцитов является наличие множества впячиваний (кавеол) плазмолеммы, содержащих ионы кальция.
Секреторные миоциты (синтетические) по своей ультраструктуре напоминают фибробласты, однако содержат в цитоплазме пучки тонких миофиламентов, расположенные на периферии клетки. В цитоплазме хорошо развиты комплекс Гольджи, гранулярная эндоплазматическая сеть, много митохондрий, гранул гликогена, свободных рибосом и полисом. По степени зрелости такие клетки относят к малодифференцированным.
Сократительный аппарат миоцитов представлен тонкими актиновыми филамен-тами (гладкомышечным альфа-актином), связанными с тропомиозином. Толстые нити состоят из миозина, мономеры которого располагаются вблизи филаментов актина. Соотношение актиновых и миозиновых филаментов в гладком миоците составляет 12 к 1. Важным компонентом контрактильного аппарата миоцитов являются электронно-плотные структуры — тельца прикрепления, расположенные свободно в цитоплазме (плотные тельца) или тесно связанные с плазмолеммой. Основными белковыми компонентами плотных телец являются альфа-актинин, актин (немышечный) и кальпонин, что позволяет расссматривать их как функциональный эквивалент Z-линий миофибрилл скелетной мышцы. Актиновые филаменты фиксируются на плотных тельцах. Промежуточные филаменты, включающие десмин и виментин, обеспечивают связи между плотными тельцами и плазмолеммой, образуя прикрепительные пластины.
Сократительные белки формируют решетчатую структуру, закрепленную по окружности плазмолеммы, поэтому сокращение выражается в укорочении клетки, которая приобретает складчатую форму, тогда как в состоянии покоя клетка вытянута. При возникновении нервного импульса, распространяющегося по плазмолемме миоцита, происходит повышение уровня внутриклеточного Са2+, который поступает в цитоплазму из кавеол, отшнуровывающихся в цитоплазму в виде пузырьков. Высвобождение ионов кальция приводит к каскаду реакций, в результате которого происходит полимеризация миозина и образование перекрестных связей миозина вдоль актиновых филаментов по мере развития мышечного сокращения. Расслабление мышцы возникает при восстановлении концентрации исходного уровня Са2+ внутри клетки путем его перемещения внутрь саркоплазматической сети. При этом образовавшиеся в присутствии ионов кальция связи между актином и миозином нарушаются, акто-миозиновый комплекс распадается, гладкий миоцит расслабляется.
Гладкие миоциты синтезируют протеогликаны, гликопротеиды, проколлаген, проэластин, из которых формируются коллагеновые и эластические волокна и основное вещество межклеточного матрикса.
Взаимодействие миоцитов осуществляется с помощью цитоплазматических мостиков, взаимных впячиваний, нексусов, десмосом или простых участков мембранных контактов клеточных поверхностей.
Регенерация гладкой мышечной ткани
Гладкая мышечная ткань висцерального и сосудистого видов обладает значительной чувствительностью к воздействию экстремальных факторов.
В активированных миоцитах возрастает уровень биосинтетических процессов, морфологическим выражением которых являются синтез сократительных белков, укрупнение и гиперхроматоз ядра, гипертрофия ядрышка, возрастание показателей ядерно-цитоплазменного отношения, увеличение количества свободных рибосом и полисом, активация ферментов, аэробного и анаэробного фосфорилирования, мембранного транспорта. Клеточная регенерация осуществляется как за счет дифференцированных клеток, обладающих способностью вступать в митотический цикл, так и за счет активизации камбиальных элементов (миоцитов малого объема).
Читайте также: Оксфорд ткань для юбки
При действии ряда повреждающих факторов отмечается фенотипическая трансформация контрактильных миоцитов в секреторные. Данная трансформация часто наблюдается при повреждении интимы сосудов, формировании ее гиперплазии при развитии атеросклероза.
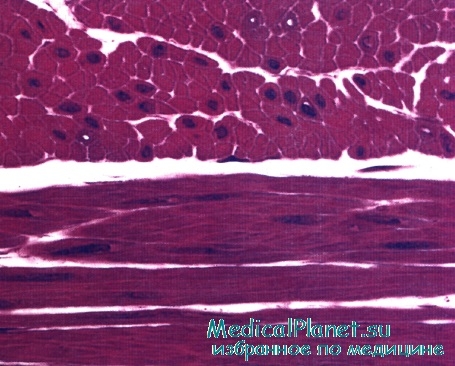
Гладкая мышечная ткань в поперечном (наверху) и продольном (внизу) разрезах. Обратите внимание на центрально расположенные ядра. Во многих клетках ядра не попали в срез.
Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
Мышечная ткань нейрального происхождения
Миоциты этой ткани развиваются из клеток нейрального зачатка в составе внутренней стенки глазного бокала. Тела этих клеток располагаются в эпителии задней поверхности радужки. Каждая из них имеет отросток, который направляется в толщу радужки и ложится параллельно ее поверхности. В отростке находится сократительный аппарат, организованный так же, как и во всех гладких миоцитах. В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы: суживающую и расширяющую зрачок.

Целомические поперечнополосатые мышечные ткани

Целомические поперечнополосатые мышечные ткани представлены у многоклеточных целомическими мышцами, возникавшими у вторичноротых и первичноротых (у последних, по-видимому, многократно) независимо. У хордовых и головоногих моллюсков такие мышцы формируют сократительный аппарат стенки сердца. У членистоногих помимо стенки трубчатых сердец целомическая мышечная ткань образует еще и стенку кишки. Несмотря на независимое происхождение целомической поперечнополосатой мышечной ткани у хордовых, головоногих моллюсков и членистоногих, она везде имеет принципиально сходное строение на тканевом уровне.
У позвоночных, в частности у млекопитающих, целомическая мышечная ткань сформирована анастомозирующими клеточными волокнами, создающими целостную в функциональном и структурном отношениях сократительную систему. Образуются такие волокна не путем слияния клеток, как скелетные симпластические мышечные волокна, а клетками-миоцитами. Волокно сердечной мышцы образуется объединением клеток специализированными межклеточными контактами.
Миотомные поперечнополосатые мышечные ткани
Источником развития элементов скелетной (соматической) поперечнополосатой мышечной ткани являются клетки миотомов — миобласты. Одни из них дифференцируются на месте и участвуют в образовании так называемых аутохтонных мышц. Другие клетки мигрируют из миотомов в мезенхиму. Они уже детерминированы, хотя внешне не отличаются от других клеток мезенхимы. Их дифференцировка продолжается в местах закладки других мышц тела.

В ходе дифференцировки возникают две клеточные линии. Клетки одной из линий сливаются, образуя удлиненные симпласты — мышечные трубочки (миотубы). В них происходит дифференцировка специальных органелл — миофибрилл. В это время в миотубах отмечается хорошо развитая гранулярная эндоплазматическая сеть. Миофибриллы сначала располагаются под плазмолеммой, а затем заполняют большую часть миотубы. Ядра, напротив, из центральных отделов смещаются к периферии. Клеточные центры и микротрубочки при этом полностью исчезают. Гранулярная эндоплазматическая сеть редуцируется в значительной степени. Такие дефинитивные структуры называют миосимпластами.
Клетки другой линии остаются самостоятельными и дифференцируются в миосателлитоциты (или миосателлиты). Эти клетки располагаются на поверхности миосимпластов.
Мышечная ткань мезенхимного типа в составе органов
Миоциты объединяются в пучки, между которыми располагаются тонкие прослойки соединительной ткани. В эти прослойки вплетаются ретикулярные и эластические волокна, окружающие миоциты. В прослойках проходят кровеносные сосуды и нервные волокна. Терминали последних оканчиваются не непосредственно на миоцитах, а между ними. Поэтому после поступления нервного импульса медиатор распространяется диффузно, возбуждая сразу многие клетки. Гладкая мышечная ткань мезенхимного происхождения представлена главным образом в стенках кровеносных сосудов и многих трубчатых внутренних органов, а также образует отдельные-мелкие мышцы (цилпарные).
Гладкая мышечная ткань в составе конкретных органов имеет неодинаковые функциональные свойства. Это обусловлено тем, что на поверхности органов имеются разные рецепторы к конкретным биологически активным веществам. Поэтому и на многие лекарственные препараты их реакция неодинакова. Возможно, разные функциональные свойства тканей связаны и с конкретной молекулярной организацией актиновых филаментов.
Мышечная ткань эпидермального происхождения
Миоэпителиальные клетки развиваются из эпидермального зачатка. Они встречаются в потовых, молочных, слюнных и слезных железах и имеют общих предшественников с их секреторными клетками. Миоэпителиальные клетки непосредственно прилежат к собственно эпителиальным и имеют общую с ними базальную мембрану. При регенерации те и другие клетки тоже восстанавливаются из общих малодифференцированных предшественников. Большинство миоэпителиальных клеток имеют звездчатую форму. Эти клетки нередко называют корзинчатыми: их отростки охватывают концевые отделы и мелкие протоки желез. В теле клетки располагаются ядро и органеллы общего значения, а в отростках — сократительный аппарат, организованный, как и в клетках мышечной ткани мезенхимного типа.
Читайте также: Толщина нитей ткани это
Мышечная ткань нейрального происхождения
Миоциты этой ткани развиваются из клеток нейрального зачатка в составе внутренней стенки глазного бокала. Тела этих клеток располагаются в эпителии задней поверхности радужки. Каждая из них имеет отросток, который направляется в толщу радужки и ложится параллельно ее поверхности. В отростке находится сократительный аппарат, организованный так же, как и во всех гладких миоцитах. В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы — суживающую и расширяющую зрачок.
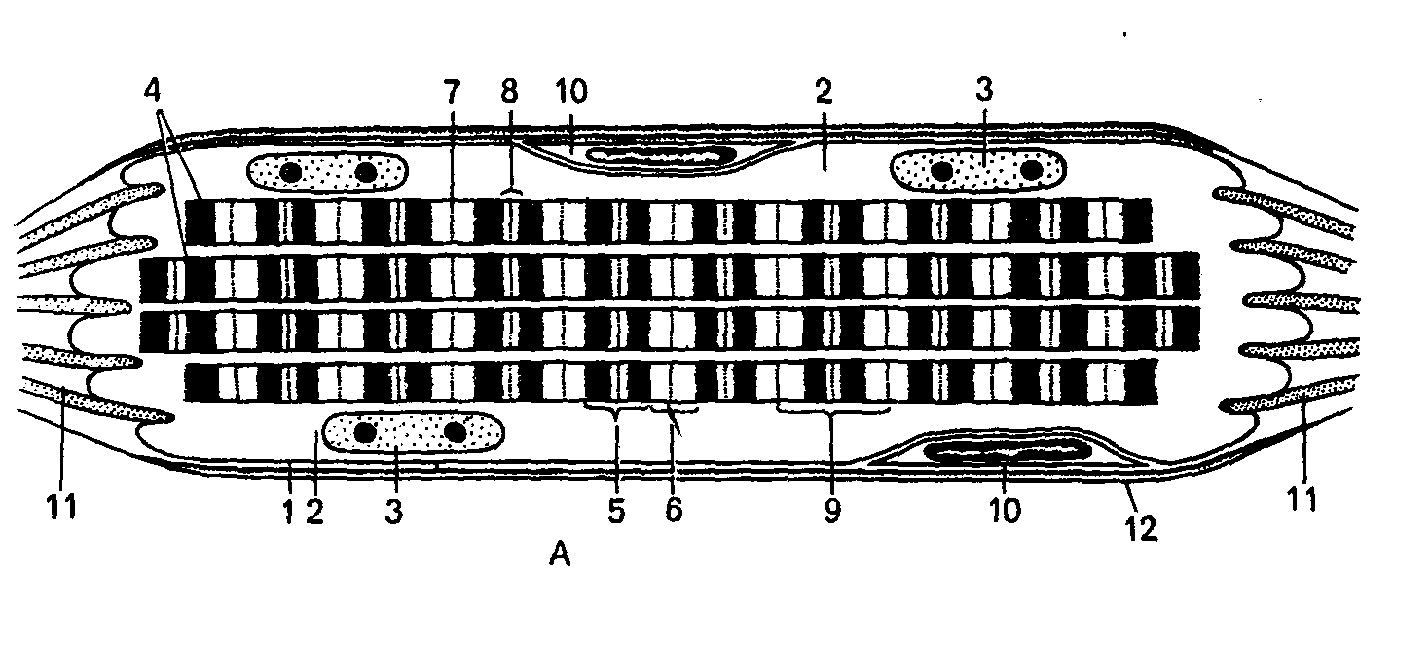
Рис 1. Поперечнополосатое мышечное волокно (миосимпласт).
I — строение мышечного волокна на светооптическом уровне. А — схема, Б — микрофотография. 1 — плазмолемма; 2 — саркоплазма; 3 — ядра миосимпласта;
5 — анизотропный диск (полоска А);
6 — изотропный диск (полоска I);
7 — телофрагма (линия Z); 8 — светлая зона (полоса Н), в середине которой проходит мезофрагма (линия М); 9 — саркомер; 10 — миосателлитоцит; 11 — сухожильные волокна; 12 — базальная мембрана
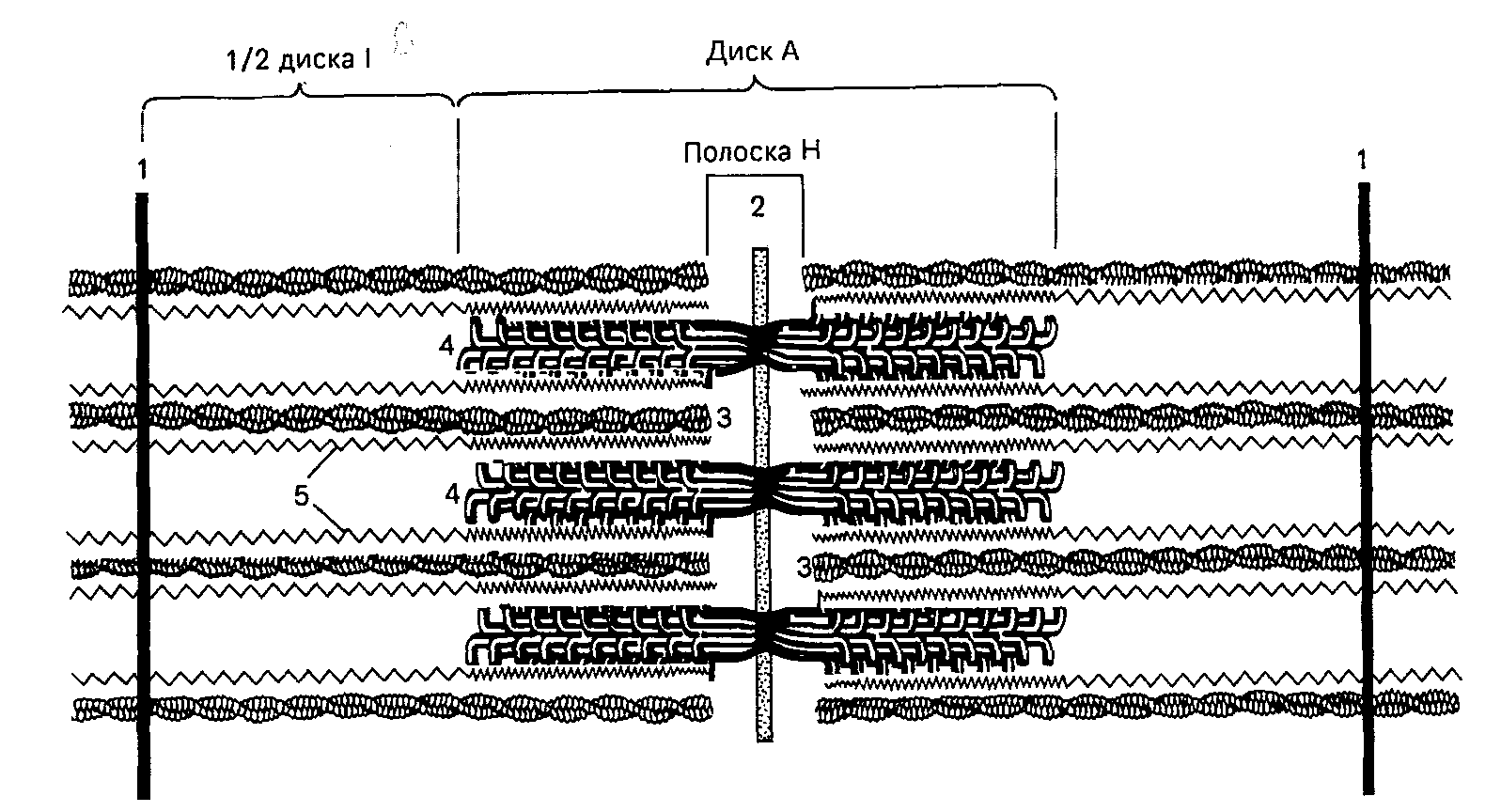
1 — линия Z; 2 — линия М; 3 — филаменты актина; 4 — филаменты миозина; 5 — фибриллярные молекулы титина

Рис. 127. Строение гладкой (неисчерченной) мышечной ткани (объемная схема) (по Р.Кристичу, с изменениями).
1 — веретеновидные гладкие миоциты; 2 — цитоплазма миоцита; 3 — ядра миоцитов; 4 — плазмолемма; 5 — базальная мембрана; 6 — поверхностные пиноцитозные пузырьки; 7 — межклеточные соединения; 8 — нервное окончание;
9 — коллагеновые фибриллы; 10 -микрофиламенты.
Мышечная ткань нейрального происхождения
Миоциты этой ткани развиваются из клеток нейрального зачатка в
составе внутренней стенки глазного бокала. Тела этих клеток располагаются в
эпителии задней поверхности радужки. Каждая из них имеет отросток,
который направляется в толщу радужки и ложится параллельно ее
поверхности. В отростке находится сократительный аппарат, организованный так же, как и во всех гладких миоцитах. В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы — суживающую и расширяющую зрачок.
Гистогенез гмт
ГМТ в эмбриональном периоде развивается из мезенхимы. Вначале мезенхимные клетки имеют звездчатую, отросчатую форму, а при дифференцировке в ГМ-клетки приобретают веретеновидную, овальную форму – миобласты (способны к размножению); в цитоплазме накапливаются органоиды спецназначения — миофибриллы из актина и миозина.
Регенерация гмт
1. Митоз миоцитов после дедифференцировки: миоциты утрачивают сократительные белки, исчезают митохондрии и превращаются в миобласты. Миобласты начинают размножаться, а потом вновь дифференцируются в зрелые леомиоциты.
2. Возможно образование новых ГМ-клеток из малодифференцированных стволовых клеток фибробластического дифферона рыхлой с.д.т.
Поперечно-полосатая МТ соматического типа (скелетная мускулатура)- является древнейшей гистологической системой. В эмбриогенезе ПП МТ соматического типа развивается из миотомов.
Структурно-функциональной единицей является мышечное волокно, состоящее из миосимпласта и миосателлитоцитов, покрытых общей базальной мембраной (длина – саниметры, при толщине 50 — 100 мкм), мион = мыш. волокно + капилляры + нервные окончания.
Камбий. Миосателлитоциты — малодифференцированные клетки, являющиеся источником регенерации мышечной ткани. Они прилежат к поверхности миосимпласта, так что их плазмолеммы соприкасаются.
Миосимпластическая часть. Мышечное волокно по форме организации живого вещества является симпластом (огромная масса цитоплазмы, где разбросаны сотни тысяч ядер).
Мышечное волокно включает большое число ядер, саркоплазму. В саркоплазме находятся:
— органоиды спецназначения — миофибриллы
— Т-система (Т-трубочки, Л-трубочки, цистерны;)
— включенияя (особенно гликоген).
Комплекс, состоящий из плазмолеммы миосимпласта и базальной мембраны, называют сарколеммой.
Строение миосимпласта. Миосимпласт имеет множество продолговатых ядер, расположенных непосредственно под сарколеммой. Их количество в одном симпласте может достигать нескольких десятков тысяч. У полюсов ядер располагаются органеллы общего значения — аппарат Гольджи и небольшие фрагменты гранулярной эндоплазматической сети. Миофибриллы заполняют основную часть миосимпласта и расположены продольно.
Саркомер — структурная единица миофибриллы, это участок между двумя соседними телофрагмами.. Каждая миофибрилла имеет поперечные темные диски (анизотропные, полоска А, представлена толстыми миозиновыми нитями) и светлые диски (изотропные, полоски I, представлена тонкими актиновыми нитями), имеющие неодинаковое лучепреломление. Каждая миофибрилла окружена продольно расположенными и анастомозирующи- ми между собой петлями агранулярной эндоплазматической сети — саркоплазматической сети. Соседние саркомеры имеют общую пограничную структуру — Z-линию (по центру светлых И-дисков (телофрагма). По центру темных А-дисков проходит светлая зона (полоса Н), в середине которой проходит мсзофрагма (линия М).
Саркомер = ½ полоски I (актин.) – полоска А (миозин.) – ½ полоски I (актин.)

5 — анизотропный диск (полоска А);
6 — изотропный диск (полоска I);
8 — светлая зона (полоса Н), в середине которой проходит мсзофрагма (линия М):
Читайте также: Как определить ткань штапель
10 — миосател- литоцит; II — сухожильные волокна;
12 — базальная мембрана (по А.Н.Студитскому).
Кроме сократительных белков актина и миозина в саркоплазме имеются еще вспомогательные белки — Тропонин и трпомиозин — они участвуют при обеспечении (поставке) сократительных белков ионами кальция, являющихся катализатором при взаимодействии актина и миозина.
При сокращении между актиновыми и миозиновыми протофибриллами при наличии катализатора — ионов кальция образуются мостики или акто-миозиновые комплексы и это обеспечивает скольжение нитей навстречу друг к другу и укорочение саркомеров.
Канальцы саркоплазматического ретикулума располагаются в продольном направлении и образуют L-трубочки (longentidunalis = продольные); они соединяются трубочками идущими в поперечном направлении в мышечном волокне — Т-трубочками (transversus=поперечно). L- и Т-трубочки соединяются с цистернами — это своеобразные емкости для ионов кальция. В стенках цистерн имеются кальциевые насосы, откачивающие ионы Са +2 из саркоплазмы в цистерны. Нервный импульс в моторных бляшках переходит на сарколемму мышечного волокна, дальше по Т-трубочкам волна деполяризации проникает внутрь волокна, распространяется по L-трубочкам и наконец волна деполяризации проходит по стенке цистерн. В момент прохождения волны деполяризации по мембране цистерны у последней повышается проницаемость для ионов Са +2 , и кальций выбрасывается в саркоплазму и подхватывается вспомогательными белками тропонином и тропомиозином и подносится к акто-миозиновому комплексу и при наличии АТФ происходит сокращение саркомера. Кальциевый насос быстро откачивает кальций обратно в цистерны — актомиозиновый комплекс распадается, поэтому происходит расслабление мышцы. Поступление нового импульса приводит к повторению всего цикла.
По строению и функциональным особенностям выделяют мышечные волокна I типа (красные м.в.), которые содержат много митохондрий, миоглобина (придает красный цвет), высокую активность фермента сукцинатдегидрогеназы, но мало миофибрилл. Красные м.в. добывают энергию для сокращения путем аэробного оксиления гликогена, т.е. нуждаются в дыхании.
М.В. II типа (белые м.в.) содержат больше миофибрилл и относительно больше гликогена, зато меньше митохондрий и у них низка активность сукцинатдегидрогеназы. Белые м.в. энергию для сокращений получают путем анаэробного окисления гликогена, т.е. в дыхании не нуждаются.
Особо следует отметить так называемые клетки миосателлитоциты (МСЦ). МСЦ были обнаружены с помощью электронного микроскопа в 1961 году. С тех пор гистогенез и регенерация скелетной МТ рассматривается в связи с этим и МСЦ. Особенностью локализации МСЦ является то, что они располагаются между базальной пластинкой и сарколеммой м. волокна. В обычных условиях эти клетки имеют неольшие размеры (20-30 мкм в длину), палочковидное ядро с большим содержанием гетерохроматина, узкую цитоплазму окружающее ядро; органеллы представлены очень бедно. Актиновые и миозиновые протофибриллы в МСЦ не обнаруживаются. Физиологическая и репаративная регенерация ПП МТ соматического типа осуществляется за счет малодифференцированных элементов — МСЦ. При травме или большой физической нагрузке клетки МСЦ постепенно выходят из состава м.волокна, начинают делиться митозом и формируют популяцию миобластов. В последующем миобласты выстраиваются в «цепочку» и начинают сливаясь образовывать миотубулы — симпласт. Миотубулы в цитоплазме накапливают миофибриллы, митохондрии и превращаются в новые мыщечные волокна, которые включают в свой состав и симпластический компонент и резервные клетки — МСЦ.
Возрастные изменения поперечно-полосатой МТ соматического типа сопровождаются атрофией м.в., т.е. уменьшением количества и толщины миофибрилл, накоплением липофусцина и жировых включений в саркоплазме, значительным утолщением базальной мембраны вокруг сарколеммы.
Гистогенез. Источником развития элементов скелетной (соматической) оперечнополосатой мышечной ткани (textus muscularis striatus sceletalis) являются клетки миотомов — миобласты. Одни из них дифференцируются на месте и участвуют в образовании так называемых аутохтонных мышц. Другие клетки мигрируют из миотомов в мезенхиму. Они уже детерминированы, хотя внешне не отличаются от других клеток мезенхимы. Их дифференцировка продолжается в местах закладки других мышц тела. В ходе дифференцировки возникают две клеточные линии. Клетки одной из линий сливаются, образуя удлиненные симпласты — мышечные трубочки (миотубы). В них происходит дифференцировка специальных органелл — миофибрилл. В это время в миотубах отмечается хорошо развитая гранулярная эндоплазматическая сеть. Миофибриллы сначала располагаются под плазмолеммой, а затем заполняют большую часть миотубы. Ядра, напротив, из центральных отделов смещаются к периферии. Клеточные центры и микротрубочки при этом полностью исчезают. Гранулярная эндоплазматическая сеть редуцируется в значительной степени. Такие дефинитивные структуры называют миосимпластами.
Клетки другой линии остаются самостоятельными и дифференцируются в миосателлитоциты (миосателлиты). Эти клетки располагаются на поверхности миосимпластов.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом