
Мышечные ткани — это ткани, для которых способность к сокращению является главным свойством. Мышечные ткани составляют активную часть опорно-двигательного аппарата (пассивной частью являются кости, соединения костей).
Общими свойствами всех мышечных тканей является сократимость и возбудимость. К данной группе тканей относятся гладкая, поперечнополосатая скелетная и поперечнополосатая сердечная мышечные ткани. Клетки мышечной ткани имеют хорошо развитый цитоскелет, содержат много митохондрий.

Гладкая (висцеральная) мускулатура
Эта мышечная ткань встречается в стенках внутренних органах (бронхи, кишечник, желудок, мочевой пузырь), в стенках сосудов, протоках желез. Эволюционно является наиболее древним видом мускулатуры.
Состоит из веретенообразных миоцитов — коротких одноядерных клеток. Между клетками имеются межклеточные контакты — нексусы (лат. nexus — связь). Благодаря нексусам возбуждение, возникшее в одной клетке, волнообразно распространяется на все остальные клетки.

Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы внутренних органов (к примеру мочевого пузыря), сокращается медленно, практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает — сокращается и утомляется быстро.
Осуществляется сокращение с помощью клеточных органоидов — миофиламентов, которые расположены в клетке хаотично и не имеют такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их тоже изучим).
Особо заметим, что в гладкой мышечной ткани миофиламенты собираются в миофибриллы только во время сокращения. У таких временных миофибрилл не может быть регулярной организации, а значит ни у таких миофибрилл, ни у гладких миоцитов не может быть поперечной исчерченности.
Гладкая мышечная ткань сокращается непроизвольно (неподвластна воле человека). Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой. К примеру невозможно по желанию сузить или расширить бронхи, кровеносные сосуды, зрачок.

Гладкая мышечная ткань называется неисчерченной, так как не обладает поперечной исчерченностью, характерной для поперечнополосатых скелетной и сердечной мышечных тканей.
Скелетная (поперечнополосатая) мышечная ткань
Скелетная мышечная ткань образует диафрагму (дыхательную мышцу), мускулатуру туловища, конечностей, головы, голосовых связок.
В отличие от гладкой мускулатуры, скелетная образована не отдельными одноядерными клетками, а длинными многоядерными волокнами, имеющими до 100 и более ядер — миосимпластами. Миосимпласт (греч. sim — вместе + plast — образованный) представляет совокупность слившихся клеток, имеет длину от нескольких миллиметров до нескольких сантиметров (соответствует длине мышцы).
Внутри миосимпласта находится саркоплазма, снаружи миосимпласт покрыт сарколеммой. Сократительные элементы — миофибриллы (лат. fibra — волоконце) — длинные тяжеобразные органеллы в миосимпласте (около 1400).

Характерная черта данной ткани — поперечная исчерченность, выражающаяся в равномерном чередовании светлых и темных полос на мышечном волокне. Это происходит потому, что границы саркомеров в соседних миофибриллах совпадают, вследствие чего все волокно приобретает поперечную исчерченность. Теперь самое время изучить микроскопическую основу мышцы — саркомер.
Саркомер (от греч. sarco — мясо (мышца) + mere — маленький)
Саркомер — элементарная сократительная единица поперечнополосатых мышц, структурная единица миофибриллы. В состав саркомера (и миофибриллы в целом) входят миофиламенты (лат. filamentum — нить) двух типов, которые обеспечивают сократимость мышечной ткани.
Саркомер состоит из актиновых (тонких) и миозиновых (толстых) филаментов, которые образованы главным образом белками актином и миозином. Сокращение происходит за счет взаимного перемещения миофиламентов: они тянутся навстречу друг другу, саркомер укорачивается (и мышца в целом).

Источником энергии для сокращения служат молекулы АТФ. К тому же невозможно представить сокращение мышц без участия ионов кальция: именно они связываются с тропонином, что приводит к изменению конформации тропомиозина (тропонин и тропомиозин — регуляторные белки между нитями актина), за счет чего становится возможно соединение актина и миозина. При сокращении мышц выделяется тепло (сократительный термогенез).
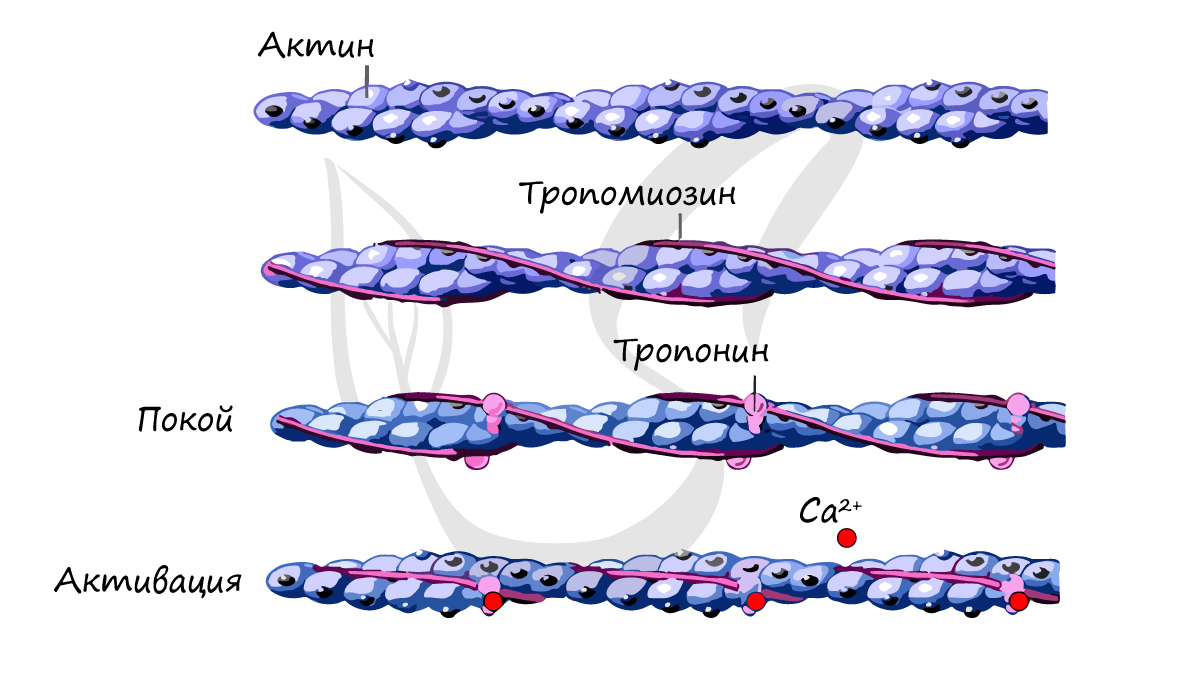
Замечу, что трупное окоченение (лат. rigor mortis) — посмертное затвердевание мышц — связано именно с ионами кальция, которые устремляются в область низкой концентрации (в саркоплазму миосимпласта), способствуя связыванию актина и миозина.
После смерти в мышце перестает синтезироваться АТФ, ее уровень быстро снижается. Как следствие этого перестает функционировать Ca-АТФаза — насос, выкачивающий ионы Ca из саркоплазмы в саркоплазматический ретикулум (мембранная органелла мышечных клеток (сходная с ЭПС), в которой запасаются ионы Ca).
В саркоплазме повышается концентрация ионов Ca — замыкаются мостики между актином и миозином, однако разомкнуться они уже не могут, в связи с чем наблюдается стойкая мышечная контрактура (лат. contractura — стягивание, сужение): конечности очень сложно разогнуть или согнуть.
Читайте также: Что можно пошить с 1 метра ткани

Вернемся к скелетным мышцам. Имеется еще ряд важных моментов, о которых нужно знать.
В процесс возбуждения вовлекается изолированно один миосимпласт, соседние миосимпласты (волокна) не возбуждают друг друга, в отличие от гладких миоцитов, где возбуждение предается между соседними клетками через нексусы. Скелетные мышцы сокращаются быстро и быстро утомляются (у гладких мышц фазы сокращения и расслабления растянуты во времени, мало утомляются) .
Скелетные мышцы сокращаются произвольно: они подконтрольны нашему сознанию. К примеру, по желанию мы можем изменить скорость движения руки, темп бега, силу прыжка. Мышцы покрыты фасцией, крепятся к костям сухожилиями, и, сокращаясь, приводят в движение суставы.

Сердечная поперечнополосатая мышечная ткань
Сердечная мышечная ткань образует мышечную оболочку сердца — миокард (от др.-греч. μῦς «мышца» + καρδία — «сердце»). Миокард — средний слой сердца, составляющий основную часть его массы. При работе сердечная мышечная ткань не утомляется.

Сердечная мышечная ткань состоит из кардиомиоцитов — одиночных клеток, имеющих поперечную исчерченность. Соединяясь друг с другом, кардиомиоциты образуют функциональные волокна.
Этот тип мышечной ткани удивительным образом сочетает свойства двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство — автоматизм.
Автоматизм — способность сердечной мышечной ткани возбуждаться и сокращаться самопроизвольно, без влияний извне. Это легко можно подтвердить, наблюдая сокращения изолированного сердца лягушки в физиологическом растворе: сокращения сердца в нем будут продолжаться несколько десятков минут после отделения сердца от организма.

Места контактов соседних кардиомиоцитов — вставочные диски (в их составе находятся нексусы), благодаря которым возбуждение одной клетки передается на соседние, таким образом волнообразно охватываются возбуждением и сокращаются новые участки миокарда.
Большое число контактов между кардиомиоцитами обеспечивает высокую эффективность и надежность проведения возбуждения по миокарду. Сокращается эта ткань непроизвольно, не утомляется.
На рисунке или микропрепарате узнать данную ткань можно по центральному положению ядер в клетках, поперечной исчерченности, наличию вставочных дисков и анастомозов (греч. anastomosis — отверстие) — мест соединений боковых поверхностей функциональных волокон (кардиомиоцитов).

В норме возбуждение проводится по проводящей системе сердца от предсердий к желудочкам (однонаправленно). Участок сердечной мышцы, в котором генерируются импульсы, определяющие частоту сердечных сокращений — водитель сердечного ритма.
Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных (англ. pacemaker — задающий ритм) клеток, которые также называют водителями ритма. Они спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.
Ответ мышц на физическую нагрузку
Физические нагрузки приводят к гипертрофии мышц (от др.-греч. ὑπερ- чрез, слишком + τροφή — еда, пища) — в них увеличивается количество мышечных волокон, объем мышечной массы нарастает.

В условиях гиподинамии (от греч. ὑπό — под и δύνᾰμις — сила), то есть пониженной активности, мышцы уменьшаются вплоть до полной атрофии (греч. а – «не» + trophe – питание). В худшем случае волокна мышечной ткани перерождаются в соединительную ткань, после чего пациент становится обездвиженным.

Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление. Гипертрофия сердца — состояние, требующее вмешательства врача и наблюдения за пациентом.
В большинстве случае гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы).

Происхождение мышц
Мышцы развиваются из среднего зародышевого листка — мезодермы.

© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
9. Особенности возбудимости и проводимости в гладких мышцах. Автоматия гладких мышц, её механизм.
Мышечные клетки соединены между собой особыми цитоплазматическими выростами – нексусами. Поэтому возбуждение в гладких мышцах легко передается с одной клетки на другую. В гладких мышцах плохо выражена проводящая возбуждение система: слабо развиты поперечные трубочки, саркоплазматический ретикулюм практически отсутствует. Поэтому скорость проведения возбуждения в гладких мышцах значительно меньше, чем в поперечно-полосатых. В гладких мышцах фактически нет синапсов, т. к. отсутствует постсинаптическая мембрана и медиатор изливается прямо на мембрану мышечных клеток, на которой находятся многочисленные рецепторы (адрено-, холино-, серотонинорецепторы). Как известно, адренорецепторы, в свою очередь, делятся на альфа и бета, поэтому взаимодействие одного и того же медиатора в зависимости от характера рецептора может давать противоположный эффект: в одном случае – сокращение гладкой мускулатуры, в другом – ее расслабление. Взаимодействие медиатора со специфическим рецептором, вызывающее тот или иной мышечный эффект опосредовано клеточными системами регуляции (цАМФ, цГМФ, Са-система). В большинстве случаев альфа-адренорецепторы участвуют в сокращении гладких мышц, в то время как бета-адренорецепторы расслабляют их. Альфа-адренорецепторы регулируют в основном уровень ионов Са, тогда как бета-адренорецепторы ингибируют аденилатциклазу. Мембрана гладких мышечных клеток не имеет Nа — каналов, в ней имеются Са — каналы, обусловливающие возбуждение благодаря возникновению Са — тока. Проводимость гладких мышц также отличается от скелетных. Как было отмечено раньше, возбуждение с одной мышечной клетки может передаваться на другую через особые контакты — нексусы, которые имеются между плазматическими мембранами соседних клеток. Возбуждение по гладким мышечным клеткам распространяется с небольшой скоростью — 2-10 см/с. Сократимость гладкой мускулатуры носит более медленный и длительный характер. Гладкие мышцы работают как в фазном, так и тоническом режиме. Вследствие большой продолжительности сократительного акта гладкие мышцы даже под влиянием редких импульсов могут переходить в состояние длительного сокращения, напоминающего тетанус. Кроме того, характерным для гладких мышц является и то, что они могут поддерживать состояние тонического напряжения без видимой затраты энергии, с чем связано их медленное утомление. Автоматия — это свойство мышечной гладкой ткани самовозбуждаться без воздействия к каких — либо факторов, которым не обладают скелетные мышцы. Клетки, обладающие автоматией, пейсмеккерные клетки, идентичны по строению обычным мышечным клеткам, однако отличаются от них по некоторым электрофизиологическим свойствам. В пейсмеккерных клетках наблюдается спонтанная деполяризация, приводящая к самовозбуждению клетки, природа которой точно не установлена. Спонтанная деполяризация, доходя до критического уровня, приводит к развитию потенциала действия и самовозбуждению мышечной клетки.
Читайте также: Мягкая полка из ткани
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
Оставленные комментарии видны всем.
Для продолжения скачивания необходимо пройти капчу:
АВТОМАТИЯ
АВТОМАТИЯ (греч. automatos — самопроизвольный, спонтанный) — способность органов или клеток к ритмической деятельности вне очевидной связи с внешними побудительными причинами. Так, например, сердце, извлеченное из организма, продолжает сокращаться в присущем ему ритме, что указывает на эндогенную (свойственную самому органу) природу АВТОМАТИИ.
В физиологии различают ритмику реактивную и ритмику автоматическую.
Ножное сердце паука-сенокосца обладает реактивной ритмикой и сокращается ритмически, но не обладает АВТОМАТИЕЙ, поскольку для каждого сокращения нужен поток импульсов. В отличие от сердца, обладающего АВТОМАТИЕЙ, оно сейчас же остановится, если его вырезать из организма. Однако в биологических процессах истинная АВТОМАТИЯ и реактивная ритмика могут быть тесно связаны между собой (например, в ритмической деятельности дыхательного центра), поэтому их следует анализировать в функциональном взаимодействии.
Наиболее ярко выраженной АВТОМАТИЕЙ обладают мышечные элементы сердца, гладкая мускулатура (кишечника, мочеточника и других органов) и нервные образования, способные к так называемой спонтанной ритмической деятельности, то есть к деятельности, не обусловленной афферентными импульсными воздействиями.
Способность к АВТОМАТИИ может быть различной у клеточных элементов одного и того же органа. Так, например, в сердце наибольшей способностью к АВТОМАТИИ обладает специфическая ткань проводящей системы в области первичных и вторичных узлов А. и в меньшей степени ткань ушков предсердий, самих предсердий и желудочков. Поэтому следует говорить о градиенте АВТОМАТИИ, то есть разной ее выраженности.
Автоматия некоторых органов или клеток может проявляться только при определенной кислотности, ионном составе, температуре, степени растяжения ткани и т. д. Так, потенциальная А. мускулатуры верхушки желудочка сердца проявляется при повышенном давлении, у гладкомышечных элементов сосудистой стенки — при растяжении и перенакоплении углекислоты и Т. д.
Читайте также: Вуаль какая плотность ткани
Изменяя условия внешней среды, можно выявить АВТОМАТИЮ в органах или тканях, которые в норме не обладают автоматической деятельностью. Например, если скелетную мышцу поместить в раствор Бидерманна, уменьшающий в мышце содержание ионов кальция, то могут появиться ритмические спонтанные подергивания отдельных волокон и даже всей мышцы в целом. Однако выявление А. в измененных условиях не доказывает, что потенциальная и нормальная А. всегда имеют одинаковые механизмы осуществления. Это важно иметь в виду при оценке опытов по исследованию механизма АВТОМАТИИ. Наличие веществ, искусственно вызывающих автоматическую активность клеточных элементов, еще не дает прямых доказательств, что естественный процесс АВТОМАТИИ происходит именно этим путем.
Проблема природы АВТОМАТИИ включает вопрос о субстрате автоматии и о клеточно-молекулярных механизмах автоматического возникновения возбуждения.
Вопрос о субстрате АВТОМАТИИ крайне сложен. У позвоночных и многих беспозвоночных животных этим субстратом являются специализированные мышечные элементы сердца, у некоторых беспозвоночных животных — разнообразные нервные клетки. К автоматической ритмической деятельности способны многие клетки центральной нервной системы. Дискуссионным остается вопрос о том, можно ли субстратом АВТОМАТИИ считать одиночные клетки или им является сложно организованный клеточный комплекс. Способность одиночных нервных и мышечных клеток сердца в культуре ткани к ритмической деятельности еще не является категорическим указанием на то, что нормальная А. нервных центров и узлов в сердце идентична потенциальной А. одиночных клеточных элементов. Проявление АВТОМАТИИ в естественных условиях всегда наблюдается в гетерогенных многоклеточных структурах, что заставляет думать о взаимодействии клеточных элементов, входящих в релаксационную систему.
Механизмы возникновения АВТОМАТИИ могут быть подразделены на эндогенно-экзогенные и системные, отражающие органоспецифическую структуру субстрата А.: нервные или мышечные. Согласно одним взглядам ритмическое возникновение возбуждения в клетках связано с фазным накоплением раздражающих веществ, определенная концентрация которых является причиной самовозбуждения клетки. Такими веществами могут быть ионы калия [Цваардемакер (Н. Zwaardemakcr)], комплекс гормонов [Хаберландт (L. Haberlandt)], ацетилхолин [Берн(J. Burn)) и т. д. Согласно другим взглядам причиной АВТОМАТИИ является фазное изменение возбудимости, позволяющее клетке не реагировать на воздействие раздражителей окружающей среды (ионы, гормоны и медиаторы) в рефрактерный период и возбуждаться от слабых постоянно действующих влияний в экзальтационный период (Д. Н. Насонов и др.).
Электрофизиологическими исследованиями установлено, что клеточные элементы, способные к автоматической ритмике, имеют сниженный трансмембранный потенциал и характерную медленную деполяризацию в интервале между двумя возбуждениями (см. Пейсмекер). Когда медленная деполяризация доходит до порогового значения, начинается быстрый процесс деполяризации как при естественном возбуждении, так и при искусственном раздражении. Медленная деполяризация в интервале между двумя возбуждениями вызвана значительным усилением в клетке натриевого тока при одновременном ослаблении калиевого.
Фазное изменение мембранной проницаемости, связанное с токами калия и натрия, раскрывает механизм автоматической ритмики, но, вероятно, еще не вскрывает ее первопричины. Естественно возникает вопрос о том, почему ритмически происходит медленная деполяризация с изменением ионных токов, напоминающим замедленную реакцию клетки на внешнее раздражение. Не исключена возможность, что медленная деполяризация является реакцией на электрическое поле, в котором находится данная клетка (эфаптическое воздействие). Разнородные клеточные элементы, обладающие разным метаболизмом, создают очень сложное электрическое ноле (между разными отделами сердца может быть разность потенциалов порядка 10— 20 мв), которое может вызвать медленную деполяризацию наиболее чувствительных клеточных элементов, способных к автоматической деятельности. При таком подходе субстратом А. должна быть не одиночная клетка, а сложная гетерогенная структура (мультиполь), включающая в себя наиболее реактивные клеточные элементы, способные к автоматической ритмике в условиях органоспецифического окружения, характеризующегося не только структурными и нейро-гуморальными, но и электротоническими взаимодействиями.
Библиография: Удельнов М. Г. и Сухова Г. С. Автоматия сердца, Усп. физиол, наук, т. 5, № 1, с. 82, 1974; Dudel J. и. Trautwein W. Der Meehanismus der automatischen rhythmischcn Impulsbildung der Herzmuskclfascr, Pfliigera Arch. ges. Physiol., Hd 267, S. 553, 1958, Bibliotrr.; Matsuda К. a. Hoshi T. The automaticity of the heart muscle cell, в кн.: The studies on the automaticity, p. 267, Tokyo. 1959.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом