
Структурно-функциональной единицей поперечнополосатой скелетной мышечной ткани является мышечное волокно. Волокно может достигать 12 см в длину, содержит большой объем саркоплазмы и сотни ядер. Каждое волокно покрыто сарколеммой, состоящей из двух слоев: внутреннего — плазмолеммы толщиной 8-10 нм и внешнего — базальной мембраны толщиной 30-40 нм. Между плазмолеммой и базальной мембраной имеется пространство шириной 15-25 нм. Кроме того, в базальную мембрану вплетаются ретикулярные волокна.
Значительный объем саркоплазмы занимают сократительные органеллы — миофибриллы. Каждая миофибрилла состоит из большого числа правильно чередующихся темных и светлых полос (дисков). В поляризованном свете темные диски обнаруживают двойное лучепреломление, поэтому называются анизотропными (А-дисками). Светлые диски таким свойством не обладают и называются изотропными (I-дисками). Каждая миофибрилла образована пучком параллельно идущих миофиламентов. А-диски состоят из толстых и тонких миофиламентов, а I-диски — только из тонких. Тонкие филаменты (5-8 нм) образованы белками актином, тропомиозином, тропонином, а толстые (10-12 нм) — миозином, белками М- и Н-полос и другими. Тонкие филаменты располагаются между толстыми, образуя гексагональное расположение.
Структурно-функциональной единицей миофибриллы является саркомер. Условная формула саркомера — 1/2 1-диска + А-диск + 1/2 I-диска. Линия сшивки соседних саркомеров соответствует Z-линии (телофрагме), которая состоит из белков альфа-актинина, десмина, вимен-тина. У позвоночных длина саркомера равна 2-3 мкм. Средняя часть миозинового диска, куда не доходят актиновые миофиламен-ты, более светлая и называется Н-полоской. Ее пересекает М-линия (мезофрагма), скрепляющая миозиновые нити посередине саркомера. В подмембранном слое сим-пласта обнаружены белки винкулин и спектрин, входящие в состав скелета симпласта.
Компоненты метаболической среды симпласта хорошо выражены. В гистогенезе с возрастанием степени зрелости симпластов наблюдается увеличение числа митохондрии, которые ориентируются по бокам Z-линии между миофибриллами и под сарколеммой. Гранулы гликогена, липидные капли формируют скопления между миофибриллами и под сарколеммой. Содержание миоглобина (связывающий кислород пигмент) варьирует в зависимости от образа жизни животного. Рибосомы представлены в виде полисом. Небольшое число лизосом принимают участие в процессах внутрисимпластической регенерации. Клеточный центр в симпласте отсутствует.
Саркоплазматическая сеть и Т-трубочки развиваются параллельно. Последние — это инвагинации плазмолеммы, которые опоясывают каждый саркомер. В продольном направлении вокруг каждой миофибриллы идут канальцы саркоплаз-матической сети. Так формируются продольная и поперечная системы, которые на срезах видны как триады. Триада — это комплекс, состоящий из поперечной трубочки и профилей двух цистерн саркоплазматической сети, расположенных симметрично по обе стороны от Т-трубочки. В цистернах саркоплазматической сети накапливаются ионы кальция, необходимые для сокращения миофибрилл.
В позднем онтогенезе происходит ряд ультраструктурных изменений в клетках и симпластах. Наиболее значимы — утолщение базальной мембраны, дезорганизация миофибрилл и Z-линии, возникновение скоплений митохондрий под сарколеммой, отделение миосателлитоцитов от симпласта и переход их в интерстициальное пространство. Иннервация мышечных волокон осуществляется двигательными нейронами передних рогов спинного мозга, которые формируют нервно-мышечные синапсы примерно в центральной части волокна.
Регенерация. Для успешной регенерации мышечной ткани необходимо сохранение напряжения мышцы, восстановление кровоснабжения и нервной связи. Основным источником регенерации являются миосателлитоциты. После активации последних происходит их митотическое деление, возникают миобласты, которые претерпевают дифференцировку, сливаются друг с другом и формируют симпласты. Развитие симпластов продолжается с участием размножающихся миосателлитоциов, часть которых сливается с растущими симпластами. Так формируются новые клеточно-симпластические системы — мышечные волокна.
Мышечные ткани гистология афанасьев
Это ткань энтомезенхимного происхождения, которая делится на два вида: висцеральную и сосудистую. В эмбриональном гистогенезе даже электронно-микроскопически трудно отличить мезенхимные предшественники фибробластов от гладких миоцитов. В малодифференцированных гладких миоцитах развиты гранулярная эндоплазматическая сеть, комплекс Гольджи. Тонкие филаменты ориентированы вдоль длинной оси клетки. По мере развития размеры клетки и число филаментов в цитоплазме возрастают. Постепенно объем цитоплазмы, занятый сократительными филаментами, увеличивается, расположение их становится все более упорядоченным. Пролиферативная активность гладких миоцитов в миогенезе постепенно снижается. Это происходит в результате увеличения продолжительности клеточного цикла, выхода клеток из цикла репродукции и перехода в дифференцированное состояние.
Однако и в дефинитивном состоянии в гладкой мышечной ткани клеточная регенерация в виде размножения миоцитов полностью не прекращается. Существуют данные о том, что пролиферация и дифференцировка в большей степени свойственна субпопуляции малых (по размерам) гладких миоцитов.
Строение гладкой мышечной ткани. Структура дефинитивных гладких миоцитов (лейомиоцитов), входящих в состав внутренних органов и стенки сосудов, имеет много общего, но в то же время характеризуется гетероморфией. Так, в стенках вен и артерий обнаруживаются овоидные, веретеновидные, отростчатые миоциты длиной 10-40 мкм, доходящие иногда до 140 мкм.
Гладкая мышечная ткань
Наибольшей длины гладкие миоциты достигают в стенке матки — до 500 мкм. Диаметр миоцитов колеблется от 2 до 20 мкм. В зависимости от характера внутриклеточных биосинтетических процессов различают контрактилъные и секреторные миоциты. Первые специализированы на функции сокращения, но вместе с тем сохраняют секреторную активность. Плазмолемма расслабленной клетки имеет ровную поверхность, а при сокращении становится складчатой. В центре клетки имеется палочковидное ядро, которое при сокращении клетки спиралевидно изгибается. Практически все ядра миоцитов содержат диплоидное количество ДНК. Гладкая эндоплазматическая сеть занимает примерно 2-7% объема цитоплазмы, а гранулярная сеть в контрактильных миоцитах выражена плохо. Митохондрии мелкие, сферические или овоидные, расположены у полюсов ядра. Характерной чертой гладких миоцитов является наличие множества впячиваний (кавеол) плазмолеммы, содержащих ионы кальция.
Читайте также: Рубчатая ткань полотняного переплетения четыре буквы
Секреторные миоциты (синтетические) по своей ультраструктуре напоминают фибробласты, однако содержат в цитоплазме пучки тонких миофиламентов, расположенные на периферии клетки. В цитоплазме хорошо развиты комплекс Гольджи, гранулярная эндоплазматическая сеть, много митохондрий, гранул гликогена, свободных рибосом и полисом. По степени зрелости такие клетки относят к малодифференцированным.
Сократительный аппарат миоцитов представлен тонкими актиновыми филамен-тами (гладкомышечным альфа-актином), связанными с тропомиозином. Толстые нити состоят из миозина, мономеры которого располагаются вблизи филаментов актина. Соотношение актиновых и миозиновых филаментов в гладком миоците составляет 12 к 1. Важным компонентом контрактильного аппарата миоцитов являются электронно-плотные структуры — тельца прикрепления, расположенные свободно в цитоплазме (плотные тельца) или тесно связанные с плазмолеммой. Основными белковыми компонентами плотных телец являются альфа-актинин, актин (немышечный) и кальпонин, что позволяет расссматривать их как функциональный эквивалент Z-линий миофибрилл скелетной мышцы. Актиновые филаменты фиксируются на плотных тельцах. Промежуточные филаменты, включающие десмин и виментин, обеспечивают связи между плотными тельцами и плазмолеммой, образуя прикрепительные пластины.
Сократительные белки формируют решетчатую структуру, закрепленную по окружности плазмолеммы, поэтому сокращение выражается в укорочении клетки, которая приобретает складчатую форму, тогда как в состоянии покоя клетка вытянута. При возникновении нервного импульса, распространяющегося по плазмолемме миоцита, происходит повышение уровня внутриклеточного Са2+, который поступает в цитоплазму из кавеол, отшнуровывающихся в цитоплазму в виде пузырьков. Высвобождение ионов кальция приводит к каскаду реакций, в результате которого происходит полимеризация миозина и образование перекрестных связей миозина вдоль актиновых филаментов по мере развития мышечного сокращения. Расслабление мышцы возникает при восстановлении концентрации исходного уровня Са2+ внутри клетки путем его перемещения внутрь саркоплазматической сети. При этом образовавшиеся в присутствии ионов кальция связи между актином и миозином нарушаются, акто-миозиновый комплекс распадается, гладкий миоцит расслабляется.
Гладкие миоциты синтезируют протеогликаны, гликопротеиды, проколлаген, проэластин, из которых формируются коллагеновые и эластические волокна и основное вещество межклеточного матрикса.
Взаимодействие миоцитов осуществляется с помощью цитоплазматических мостиков, взаимных впячиваний, нексусов, десмосом или простых участков мембранных контактов клеточных поверхностей.
Регенерация гладкой мышечной ткани
Гладкая мышечная ткань висцерального и сосудистого видов обладает значительной чувствительностью к воздействию экстремальных факторов.
В активированных миоцитах возрастает уровень биосинтетических процессов, морфологическим выражением которых являются синтез сократительных белков, укрупнение и гиперхроматоз ядра, гипертрофия ядрышка, возрастание показателей ядерно-цитоплазменного отношения, увеличение количества свободных рибосом и полисом, активация ферментов, аэробного и анаэробного фосфорилирования, мембранного транспорта. Клеточная регенерация осуществляется как за счет дифференцированных клеток, обладающих способностью вступать в митотический цикл, так и за счет активизации камбиальных элементов (миоцитов малого объема).
При действии ряда повреждающих факторов отмечается фенотипическая трансформация контрактильных миоцитов в секреторные. Данная трансформация часто наблюдается при повреждении интимы сосудов, формировании ее гиперплазии при развитии атеросклероза.
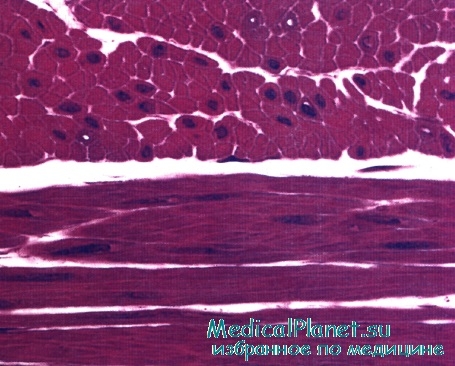
Гладкая мышечная ткань в поперечном (наверху) и продольном (внизу) разрезах. Обратите внимание на центрально расположенные ядра. Во многих клетках ядра не попали в срез.
Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
«Ростовский государственный медицинский университет»
МИНЗДРАВСОЦРАЗВИТИЯ РОССИЙСКОЙ ФЕДЕРАЦИИ
Кафедра гистологии, цитологии и эмбриологии
/под ред. д.м.н. , профессора П.А.Хлопонина/
Экскурс
содержания практического и самостоятельного занятия по
теме «Мышечные ткани» в курсе общей гистологии.
1. Классификации мышечных тканей.
2. Функции скелетной мышечной ткани.
3. Компоненты скелетного мышечного волокна.
4. Гистогенез скелетной мышечной ткани.
5. Строение сарколеммы скелетного мышечного волокна.
7. Состав саркоплазмы и органеллы общего значения миосимпласта.
8. Строение миофибриллы при световой и электронной микроскопии.
9. Промежуточные филаменты и включения миосимпласта.
10. Типы скелетных мышечных волокон.
11. Гистофизиология скелетной мышечной ткани.
12. Скелетная мышца как орган.
13. Регенерация скелетной мышечной ткани.
14. Гистогенез сердечной мышечной ткани.
15. Структурная и функциональная единица сердечной мышцы.
16. Строение вставочных дисков.
17. Типы кардиомиоцитов, сократительные кардиомиоциты.
18. Сократительно-секреторные кардиомиоциты.
19. Проводящие кардиомиоциты, их типы.
20. Гистофизиология сердечной мышечной ткани.
21. Регенерация сердечной мышечной ткани.
22. Эмбриогенез гладкой мышечной ткани.
23. Сарколемма и ядра гладкого миоцита.
24. Органеллы общего значения гладкого миоцита.
25. Сократительные и промежуточные филаменты гладкого миоцита.
26. Гистофизиология гладкой мышечной ткани.
27. Структурная и функциональная единица гладкой мышечной ткани.
28. Регенерация гладкой мышечной ткани.
1 . Морфологическая классификация. Существует три вида мышечных тканей: а) поперечнополосатая скелетная мышечная ткань; б) поперечнополосатая сердечная мышечная ткань; в) гладкая мышечная ткань.
Гистогенетическая классификация мышечных тканей включает следующие типы: 1) соматические, или миотомные (скелетная, поперечнополосатая) – развивается из миотомов сомитов; 2) целомические (сердечная) – развивается из миоэпикардиальной пластинки; 3) мезенхимные (гладкая мышечная ткань) – развивается из мезенхимы; 4) эпидермальные (миоэпителиальные клетки некоторых желез) – развиваются из эктодермы и прехордальной пластинки; 5) нейральные (мышцы радужки глаза) – являются производными нейроэктодермы.
Читайте также: Снижение объемов тела или тканей называется
Скелетная мышечная ткань
2. Скелетная мышечная ткань выполняет следующие функции:
а) движения (сокращения – расслабления); входит в качестве основной ткани в состав скелетных мышц – органов движения; б) терморегуляции; сокращение скелетной мышечной ткани ведёт к образованию большого количества тепла, что в условиях холода обеспечивает согревание тела (так называемый сократительный термогенез).
3. Скелетное мышечное волокно является структурно-функциональной единицей скелетной мышечной ткани. Каждое волокно включает: — миосимпласт – длинную цилиндрическую структуру со множеством ядер, способную к сокращению; — миосателлитоциты – мелкие одноядерные клетки в углублениях миосимпласта, играющие роль камбия; — базальную мембрану, окружающую симпласт вместе с миосателлитами.
4. Скелетная мышечная ткань развивается из миотомов мезодермальных сомитов, поэтому называется соматической. Клетки миотомов дифференцируются в двух направлениях: 1) из одних образуются миосимпласты; 2) из других — миосателлитоциты
Образование миосимпластов. Клетки миотомов дифференцируются в миобласты, которые сливаются вместе, образуя мышечные трубочки (миотубы). В последних ядра лежат вдоль оси, посередине. Последующее закономерное накопление в саркоплазме миофибрилл ведет к оттеснению ядер на периферию и к образованию функционально активных миосимпластов. Миосателлитоциты – обособившиеся в ходе миогенеза G1-миобласты.
5. Сарколемма – оболочка мышечного волокна — образована цитолеммой и базальной мембраной. Цитолемма образует тонкие впячивания вглубь мышечного волокна – поперечные трубочки (синонимы — Т-трубочки, Т-канальцы от «transversus» — поперечный); — базальная мембрана образует с цитолеммой соединения, обеспечивающие передачу усилия сокращения мышечных волокон через сухожилия на скелет (отсюда название «скелетная мышечная ткань»).
Между базальной мембраной и цитолеммой мышечных волокон расположены миосателлитоциты. Они участвуют в регенерации скелетной мышечной ткани.
6. Ядра миосимпласта — располагаются под цитолеммой, их большое количество; они имеют палочковидную форму; при резком сокращении волокон могут деформироваться и штопорообразно скручиваться; — в миосимпласте отсутствуют центриоли, отчего ядра не способны делиться.
7. Саркоплазма (цитоплазма) – содержит: а) органеллы общего значения, б) органеллы специального значения – миофибриллы, в) включения.
Органеллы общего значения такие как комплекс Гольджи, гранулярная ЭПС, свободные рибосомы, лизосомы развиты слабо и располагаются у полюсов ядер. Хорошо развиты митохондрии и гладкая ЭПС.
Гладкая ЭПС (саркоплазматическая сеть) специализирована на депонировании ионов кальция. Это система замкнутых полостей, большей частью канальцев, имеющих, преимущественно, продольное направление, называющихся L-канальцами (longitudinalis – продольные). Подойдя к Т-трубочкам, L-канальцы сливаются и образуют поперечные терминальные цистерны (Т-цистерны), проходящие рядом с Т-трубочками параллельно им. В результате образуются комплексы – триады, состоящие из двух терминальных цистерн и одной Т-трубочки. В локусах максимального сближения мембран поперечных и продольных канальцев возможно распространение возбуждения в направлении L — системы и триады играют важную роль в инициации мышечного сокращения, проявляющейся в виде перемещения ионизированного кальция.
8. Миофибриллы – органеллы специального значения мышечной ткани.
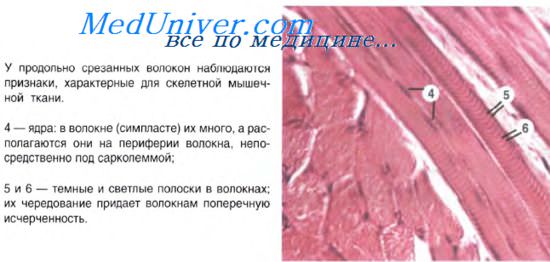
При световой микроскопии в каждой миофибрилле обнаруживается поперечная исчерченность, обусловленная наличием светлых и тёмных дисков; светлые диски называют изотропными или I-дисками; посередине I-диска проходит Z-линия (телофрагма); — тёмные диски называют анизотропными или А-дисками; в центре А-диска выявляется светлая полоска Н, а посередине её проходит тёмная линия М (мезофрагма).
Саркомер – участок миофибриллы, лежащий между двумя соседними Z-линиями. Каждый саркомер включает в себя: диск А и по сторонам от него два полудиска I (примыкающие к соседним Z-линиям). Саркомер – структурно-функциональная единица миофибриллы и всего мышечного волокна, так как в его составе есть все компоненты, участвующие в сокращении.
При электронной микроскопии миофибриллы состоят из тонких и толстых миофиламентов. Взаимное расположение их: а/ свободные концы тонких миофиламентов располагаются между концами толстых; б/ в пределах Н-зоны А-диска содержатся только толстые миофиламенты; в/ в тёмных областях А-диска – и тонкие, и толстые миофиламенты (лежащие параллельно друг другу); г/ в пределах I-диска видны только тонкие миофиламенты.
Для тонких миофиламентов (актиновых) характерны: а/ в их состав входят белки актин, тропонин и тропомиозин; б/ молекулы актина имеют гранулярное строение (G-актин) и, соединяясь вместе, образуют длинные цепи (фибриллярный F-актин); в / в актиновых филаментах таких цепей две, они образуют двойную спираль; г/ в бороздках между спиральными цепями актина лежат молекулы тропомиозина, также образуя две спирали; к молекулам тропомиозина на равных расстояниях друг от друга прикрепляются молекулы тропонина; д/ тропониновый комплекс состоит из трёх глобулярных субъединиц: TnT, TnI, TnC; TnT осуществляет прикрепление тропонинового комплекса к тропомиозину; TnC отвечает за связывание с ионами Ca 2+ ; TnI препятствует взаимодействию миозиновых головок с актином; е/ в центре тонких миофиламентов выявляется белок альфа-актинин служит местом (локусом) скрепления тонких миофиламентов, т.е. морфохимическим компонентом телофрагмы (видимой как Z-линия).
Толстые миофиламенты (миозиновые): а/ состоят, в основном, из молекул белка миозина; б/ молекула миозина похожа на клюшку для игры в гольф, в которой различают двойную «головку» (ту часть клюшки, которая ударяет по мячу) и хвостовую часть (рукоятка клюшки); в/ головка миозина имеет АТФ-азную активность и способна расщеплять АТФ с образованием энергии; хвостовая часть может сгибаться в двух местах (шарнирные участки); г/ молекулы миозина соединяются в пучки и формируют толстую нить; их головки «торчат» из стержня в периферических отделах миозиновых филаментов; в центральной части головок нет; д/ толстые миофиламенты соединены в центре мезофрагмой, являющейся опорной структурой в центре саркомера; е/ белок титин прикрепляет концы толстых миофиламентов к телофрагмам и препятствует перерастяжению миофибрилл.
Читайте также: Хорошая швейная машинка для толстых тканей
9. Промежуточные филаменты состоят из белка десмина. Благодаря им поддерживается упорядоченное взаимное расположение саркомеров миофибрилл и других компонентов мышечного волокна.
Обнаруживаемые включения гликогена и липидов используются для получения энергии. Миоглобин, выявляемый в саркоплазме скелетных мышечных волокон, является железосодержащим пигментом, аналогичным гемоглобину. Он способен связывать кислород.
10. Различают три типа скелетных мышечных волокон. I тип – красные мышечные волокна. В них преобладает саркоплазма, в которой много белка миоглобина (обеспечивающего красный цвет волокон), липидов и митохондрий, но сравнительно малый её объем, занимаемый миофибриллами и включениями гликогена. Это медленные (тонические) мышечные волокна. Они менее сильные и менее утомляемые.
II тип – белые мышечные волокна. В них больше миофибрилл и гликогена, но меньше миоглобина, липидов и митохондрий. Это быстрые (тетанические) мышечные волокна. Они могут сокращаться быстрее и с большей силой, но и быстрее утомляются. III тип скелетных мышечных волокон занимает в структурном и в функциональном отношении промежуточное положение между первыми двумя.
11. Механизм мышечного сокращения. Общепринятой является теория скользящих нитей. Этапы гистофизиологии скелетного мышечного волокна:
а/ передача проходящего по нервному волокну импульса на плазмолемму мышечного волокна и распространение его по Т-трубочкам внутрь мышечного волокна к локусам специализированных контактных взаимоотношений с лежащими рядом мембранами терминальных цистерн саркоплазматической сети;
б/ из цистерн саркоплазматической сети после их возбуждения в саркоплазму выходят ионы Са 2+ , которые связываются с молекулами тропонина (TnC);
в/ при связывании Са 2+ изменяется конфигурация тропонина, и актиновые центры, блокированные TnI, «открываются»; при этом, головки миозина, обладающие адгезивностью, приобретают возможность взаимодействовать с молекулами актина;
г/ молекулы миозина изгибаются в шарнирных областях и присоединяются к молекулам актина, создавая тянущее усилие; и этот процесс сопровождается гидролизом АТФ обеспечивающим энергетику сокращения;
д/ после этого молекулы миозина отсоединяются от активных актиновых участков и вновь присоединяются, но в новом месте, что также сопровождается гидролизом АТФ; происходит всё более глубокое вдвигание толстых миофиламентов между тонкими;
е/ в результате уменьшается длина каждого саркомера и, следовательно, всех миофибрилл, всех мышечных волокон в целом;
ё/ при отсутствии нервных импульсов Са 2+ вновь откачивается в саркоплазматическую сеть, и активные центры на актиновых филаментах закрываются тропонином; происходит расслабление миофибрилл.
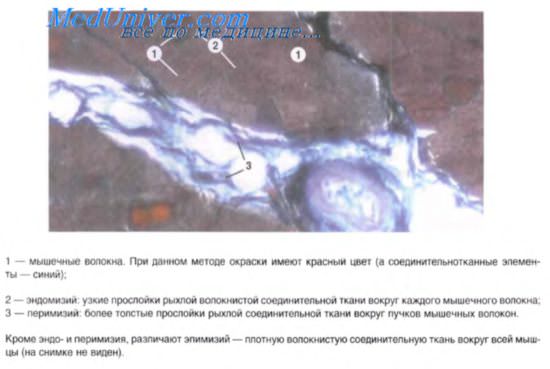
12. Мышца как орган образована: мышечными волокнами и комплексом прослоек соединительной ткани, в которых находятся сосуды и нервы.
Стромальные прослойки скелетных мышц представляют: а/ эндомизий — это тонкие прослойки рыхлой волокнистой соединительной ткани, расположенные вокруг каждого мышечного волокна и связанные через базальную мембрану сарколеммы с цитолеммой мышечных волокон; б/ перимизий – более толстые прослойки рыхлой волокнистой соединительной ткани вокруг группы мышечных волокон; в/ эпимизий – плотная оформленная соединительная ткань, окружающая всю мышцу. Эти соединительнотканные прослойки взаимосвязаны и без резкой границы переходят в сухожилие.
Мион и нервно-мышечная единица. Каждое мышечное волокно сопровождается сетью гемокапилляров и имеет собственную иннервацию; комплекс этих элементов называется мионом. Иннервация скелетной мышечной ткани осуществляется двигательными и чувствительными нервными волокнами, заканчивающимися нервными окончаниями. От миона следует отличать нервно-мышечную единицу — группу мышечных волокон, иннервируемых одним мотонейроном.
Связь мышцы с сухожилием. На концах мышечных волокон имеются впячивания сарколеммы. В эти впячивания входят коллагеновые и ретикулярные волокна сухожилий. Ретикулярные волокна прободают базальную мембрану и при помощи молекулярных сцеплений соединяются с цитолеммой. Затем эти волокна возвращаются в просвет впячивания и оплетают коллагеновые волокна сухожилия, как бы привязывая их к мышечному волокну. Коллагеновые волокна образуют сухожилия, которые прикрепляются к костному скелету.
13. Физиологическая регенерация – обновление мышечных волокон. При этом, миосателлитоциты вступают в циклы пролиферации с последующей дифференцировкой в миобласты и их включением в состав предсуществующих мышечных волокон.
Репаративная регенерация – восстановление мышечных волокон после повреждения. Первый способ – восстановление целостности повреждённого волокна за счёт медленного роста его концов (в месте разрыва) навстречу друг другу. Второй способ – образование новых мышечных волокон. При этом в морфологическом механизме этого происходят практически те же события, что и в эмбриогенезе: а/ размножение миосателлитоцитов и превращение их в миобласты; б/ слияние миобластов друг с другом, в результате чего образуются мышечные трубочки с центральным расположением ядер;
в/ накопление миофибрилл и оттеснение ядер на периферию миосимпласта.
Однако при значительном повреждении базальной мембраны мышечных волокон полного восстановления прежней структуры обычно не происходит: дефект мышцы прорастает соединительной тканью.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом