
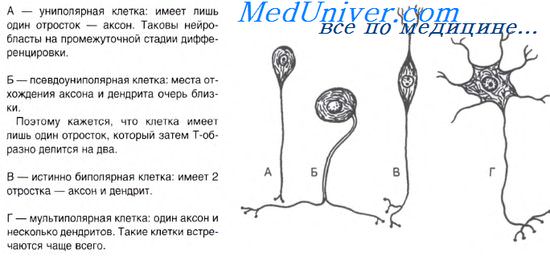
Нервная ткань в ходе гистогенеза имеет происхождение
Ткани нервной системы выполняют важнейшую функцию организма — функцию реактивности, основанную на способности нервных клеток воспринимать раздражение, вырабатывать и передавать нервные импульсы. Они участвуют в получении, хранении и переработке информации из внешней и внутренней среды организма, обеспечивают регуляцию и интеграцию деятельности всех органов и систем человека.
В каждой части нервной системы клеточный состав нервной ткани и ее морфофункциональные особенности неповторимы. Нервная ткань коры большого мозга, нервная ткань спинного мозга, нейросекреторная ткань гипоталамуса, нервная ткань ствола мозга, нервная ткань вегетативных ганглиев и других частей нервной системы — все это разновидности тканей нервной системы с достаточно четкими, специфическими (органотипическими) и стойко закрепленными признаками.
Особую группу вспомогательных тканей в нервной системе образует нейроглия, или макроглия, и ее разновидности (эпендима, астроглия, олигодендроглия и др.). Кроме того, в состав органоспецифической нервной ткани входит микроглия, представленная диффероном макрофагов. Многие авторы рассматривают нейроглию как составную часть нервной ткани, употребляя при этом термин «нервная ткань» в единственном числе.
Гистогенез нервной системы.
Источником развития нервной ткани и нейроглии является нервная пластинка 18-суточного зародыша человека. После нейруляции из нее образуются нервная трубка и ганглиозные пластинки. Кроме того, в развитии черепных нервов принимают участие плакоды — утолщения эктодермы по бокам краниальной части будущей нервной трубки. Эмбриональные зачатки состоят из малодифференцированных клеток — медуллобластов (матричных клеток). На ранних этапах гистогенеза происходит детерминация и дивергентная дифференцировка клеток, в результате чего возникают два направления их развития: нейробластическое и глиобластическое. Из мезенхимы возникают клетки микроглии ЦНС.
В нейрогистогенезе различают стадии медуллобластов, нейробласта, молодого нейрона и зрелого нейрона. Медуллобласты интенсивно делятся митозом. Стадия нейробласта характеризуется миграцией клеток, при этом необратимо блокируется способность клеток к пролиферации. В цитоплазме нейробластов определяются хорошо развитая гранулярная эпдоплазматическая сеть, комплекс Гольджи и митохондрии. Начинается синтез специфических белков нервных клеток, входящих в состав нейрофиламентов и микротрубочек. Появляется конус роста аксона.
Стадия молодого нейрона характеризуется ростом отростков, увеличением объема клетки, образованием хроматофильной субстанции и появлением первых синапсов. Дифференцировка нейробластов в нейроны происходит группами (гнездами), так что все их аксоны растут в виде пучка нервных волокон в одном направлении, образуя в дальнейшем проводящие пути и нервы.
Важной особенностью гистогенеза является запрограммированная гибель нейронов по типу апоптоза. Например, в гистогенезе спинного мозга позвоночных до 40-50% нервных клеток передних рогов гибнет после завершения пролиферативной фазы.
Самой продолжительной стадией является стадия зрелого нейрона, на протяжении которой нейрон приобретает свою окончательную форму и специфическую гистохимическую организацию. Наряду с дифференцировкой нейронов происходит все более глубокая их интеграция в составе рефлекторных дуг. Между нейронами устанавливаются многочисленные синаптические связи. Сложный характер приобретают взаимодействия между нервными и глиальными клетками.
— Вернуться в оглавление раздела «гистология»
Нервная ткань в ходе гистогенеза имеет происхождение
В развитии низших и высших позвоночных отчетливо прослеживается единая общебиологическая закономерность, выражающаяся в появлении зародышевых листков и обособлении основных зачатков органов и тканей. Процесс образования тканей из материала эмбриональных зачатков составляет суть учения о гистогенезах.
Эмбриональный гистогенез, по определению А.А. Клишова (1984), — это комплекс координированных во времени и пространстве процессов пролиферации, клеточного роста, миграции, межклеточных взаимодействий, дифференциации, детерминации, программированной гибели клеток и некоторых других. Все названные процессы в той или иной мере протекают в зародыше, начиная с самых ранних стадий его развития.
Пролиферация. Основной способ деления тканевых клеток — это митоз. По мере увеличения числа клеток возникают клеточные группы, или популяции, объединенные общностью локализации в составе зародышевых листков (эмбриональных зачатков) и обладающие сходными гистогенетическими потенциями. Клеточный цикл регулируется многочисленными вне- и внутриклеточными механизмами. К внеклеточным относятся влияния на клетку цитокинов, факторов роста, гормональных и нейрогенных стимулов. Роль внутриклеточных регуляторов играют специфические белки цитоплазмы. В течение каждого клеточного цикла существуют несколько критических точек, соответствующих переходу клетки из одного периода цикла в другой. При нарушении внутренней системы контроля клетка под влиянием собственных факторов регуляции элиминируется апоптозом, либо на некоторое время задерживается в одном из периодов цикла.
Читайте также: Ткань для костюма шута
Метод радиографического анализа клеточных циклов в различных тканях выявил особенности соотношения клеточной репродукции и дифференцировки. Например, если в тканях (кроветворные ткани, эпидермис) имеется постоянный фонд пролиферирующих клеток, за счет которых обеспечивается непрерывное возникновение новых клеток взамен погибающих, то эти ткани относятся к обновляющимся. Другие ткани, например, некоторые соединительные, характеризуются тем, что в них увеличение количества клеток происходит параллельно с их дифференцировкой, клетки в этих тканях характеризуются низкой митотической активностью. Это растущие ткани. Наконец, нервная ткань характеризуется тем, что все основные процессы репродукции заканчиваются в период эмбрионального гистогенеза (когда формируется основной запас стволовых клеток, достаточный для последующего развития ткани). Поэтому она отнесена к стабильным (стационарным) тканям. Продолжительность жизни клеток в обновляющихся, растущих и стабильных тканях разная.
Наряду с обновлением клеточной популяции, в самих клетках постоянно наблюдается обновление внутриклеточных структур (внутриклеточная физиологическая регенерация).
Клеточный рост, миграция и межклеточные взаимодействия. Рост клеток проявляется в изменении их размеров и формы. При усилении функциональной активности и внутриклеточных биосинтезов наблюдается увеличение объема клетки. Если объем клетки превышает некую норму, то говорят о ее гипертрофии, и наоборот, при снижении функциональной активности происходит уменьшение объема клетки, а при переходе некоторых нормативных параметров возникает атрофия клетки. Рост клетки не беспределен и определяется оптимальным ядерно-цитоплазменным отношением.
Важное значение для гистогенеза имеют процессы перемещения клеток. Миграция клеток наиболее характерна для периода гаструляции. Однако и в период гисто- и органогенеза происходят перемещения клеточных масс (например, смещения миобластов из миотомов в места закладки скелетных мышц; движение клеток из нервного гребня с образованием спинномозговых ганглиев и нервных сплетений, миграция гоноцитов и т. д.). Миграция осуществляется с помощью нескольких механизмов. Так, различают хемотаксис — движение клеток в направлении градиента концентрации какого-либо химического агента (перемещения спермиев к яйцеклетке, предшественников Т-лимфоцитов из костного мозга в закладку тимуса).
Гаптотаксис — механизм перемещения клеток по градиенту концентрации адгезионной молекулы (движение клеток протока пронефроса у амфибий по градиенту щелочной фосфатазы на поверхности мезодермы). Контактное ориентирование — когда в какой-либо преграде остается один канал для перемещения (описан у рыб при образовании плавников).
Контактное ингибирование — этот способ перемещения наблюдается у клеток нервного гребня. Суть способа заключается в том, что при образовании ламеллоподии одной клеткой и контакта ее с другой клеткой, ламеллоподия прекращает рост и постепенно исчезает, но в другой части мигрирующей клетки при этом формируется новая ламеллоподия.
В процессе миграции клеток важную роль играют межклеточные взаимодействия. Существует несколько механизмов такого взаимодействия (контактного и дистантного). Выделяется большая группа молекул клеточной адгезии (МКА). Так, кадгерины — это Са2+-зависимые МКА, отвечают за межклеточные контакты при образовании тканей, за формообразование и др. В молекуле кадгерина различают внеклеточный, трансмембранный и внутриклеточный домены. Например, внеклеточный домен ответственен за адгезию клеток с одинаковыми кадгеринами, а внутриклеточный — за форму клетки. Другой класс МКА — это иммуноглобулиновое суперсемейство Са2+-независимых МКА, обеспечивающих, например, адгезию аксонов к сарколемме мышечных волокон, или миграцию нейробластов вдоль радиальных глиоцитов в закладке коры большого мозга и др. Следующий класс МКА — это мембранные ферменты — гликозилтранферазы. Последние по типу «ключ-замок» соединяются с углеводными субстратами — гликозаминогликанами надмембранного комплекса клетки, осуществляя таким образом прочное сцепление клеток.
Кроме механизмов межклеточного взаимодействия, существуют механизмы взаимодействия клеток с субстратом. Они включают формирование рецепторов клетки к молекулам внеклеточного матрикса. К последним относят производные клеток, среди которых наиболее изученными адгезионными молекулами являются коллаген, фибронектин, ламинин, тенасцин и некоторые другие. Коллагены, среди которых различают несколько десятков типов, входят в состав межклеточного вещества рыхлой волокнистой соединительной ткани, базальной мембраны и пр. Фибронектин, секретируемый клетками, является связывающей молекулой между мигрирующей клеткой и межклеточным матриксом. Ламинин — компонент базальной мембраны, также связывает мигрирующие клетки с межклеточным матриксом (справедливо по отношению к эпителиоцитам и нейробластам).
Читайте также: В какие растениях отсутствуют ткани
Для осуществления связи мигрирующих клеток с межклеточным матриксом клетки формируют специфические рецепторы. К ним относятся, например, синдекан, который обеспечивает контакт эпителиоцита с базальной мембраной за счет сцепления с молекулами фибронектина и коллагена. Интегрины клеточных поверхностей связывают с внеклеточной стороны молекулы внеклеточного матрикса, а внутри клетки — белки цитоскелета (например, актиновые микрофиламенты). Так возникает связь внутри- и внеклеточных структур, что позволяет клетке использовать для перемещения собственный сократительный аппарат. Наконец, существует большая группа молекул, формирующих клеточные контакты, осуществляющие коммуникацию между клетками (щелевые контакты), механическую связь (десмосомы, плотные контакты).
Дистантные межклеточные взаимодействия осуществляются путем секреции гормонов и факторов роста (ФР). Последние — это вещества, оказывающие стимулирующее влияние на пролиферацию и дифференцировку клеток и тканей. К ним относятся, например, ФР, полученный из тромбоцитов и влияющий на переход клеток в фазу размножения (гладких миоцитов, фибробластов, глиоцитов); эпидермальный ФР — стимулирует пролиферацию эпителиоцитов, производных эктодермы; ФР фибробластов — стимулирует пролиферацию фибробластов. Особо выделяется большая группа пептидов (соматотропины, соматомедины, инсулин, лактоген), влияющих на развитие клеток плода.
Факторы, тормозящие пролиферацию и дифференцировку клеток, также принимают кооперативное участие в процессах развития клеток и тканей. К ним относятся, например, бета-интерферон и трансформирующий ФР. Последний, однако, в отношении разных клеточных типов действует по-разному: блокирует размножение многих типов эпителиоцитов, но стимулирует размножение соединительнотканных клеток.
— Вернуться в оглавление раздела «гистология»
Нервная ткань в ходе гистогенеза имеет происхождение
6. Гистогенез нервной ткани
Нервная ткань развивается из дорсальной эктодермы. У 18-дневного эмбриона человека эктодерма по средней линии спины дифференцируется и утолщается, формируя нервную пластинку, латеральные края которой приподнимаются, образуя нервные валики, а между валиками формируется нервный желобок. Передний конец нервной пластинки расширяется, образуя позднее головной мозг. Латеральные края продолжают подниматься и растут медиально, пока не встретятся и не сольются по средней линии в нервную трубку, которая отделяется от лежащей над ней эпидермальной эктодермы. Полость нервной трубки сохраняется у взрослых в виде системы желудочков головного мозга и центрального канала спинного мозга. Часть клеток нервной пластинки не входит в состав нервной трубки и эпидермальной эктодермы и образует скопления по бокам от нервной трубки, которые сливаются в рыхлый тяж, располагающийся между нервной трубкой и эпидермальной эктодермой, — нервный гребень (ганглиозная пластинка). Из нервной трубки в дальнейшем формируются нейроны и макроглия центральной нервной системы. Нервный гребень дает начало нейронам чувствительных (сенсорных) и автономных ганглиев, клеткам мягкой мозговой и паутинной оболочек мозга и некоторым видам глии : шванновским клеткам, клеткам-сателлитам ганглиев, клеткам мозгового вещества надпочечников, меланоцитам кожи, части клеток APUD-системы, сенсорным клеткам каротидных телец и др.
В формировании ганглиев V, VII, IX и X черепных нервов принимают участие, кроме нервного гребня, также нейральные (нейрогенные) плакоды , представляющие собой утолщения эктодермы по бокам формирующейся нервной трубки в краниальном отделе зародыша.
Нервная трубка на ранних стадиях эмбриогенеза представляет собой многорядный нейроэпителий, состоящий из вентрикулярных или нейроэпителиальных клеток. В дальнейшем в нервной трубке дифференцируется 4 концентрических зоны: вентрикулярная ( эпендимная ), субвентрикулярная , промежуточная (плащевая) и краевая (маргинальная).
Вентрикулярная ( эпендимная ) зона состоит из делящихся клеток цилиндрической формы. Ядро вентрикулярной клетки мигрирует в люменальный конец клетки, обращенной к центральному каналу. Клетки делятся и после деления ядра дочерних клеток также мигрируют в апикальные части образующихся клеток, где происходит репликация ДНК. Митотический цикл и цикл ядерной миграции продолжаются от 5 до 24 ч. Вентрикулярные (или матричные) клетки являются предшественниками нейронов и клеток макроглии . Микроглия развивается из другого источника (см. ниже). Предшественники глиальных клеток отличаются по присутствию глиального фибриллярного кислого белка промежуточных филаментов в делящихся клетках вентрикулярной зоны.
Читайте также: Пограничные или эпителиальные ткани
Субвентрикулярная зона состоит из клеток, утративших способность к перемещению ядер, но сохраняющих высокую пролиферативную активность. Субвентрикулярная зона существует в области спинного мозга в течение нескольких дней, но в тех областях головного мозга, где гистогенез совершается особенно интенсивно, формируются субвентрикулярные и экстравентрикулярные герминативные (камбиальные) зоны , существующие длительное время. Так, экстравентрикулярная камбиальная зона мозжечка исчезает у человека к 20 мес постнатального онтогенеза.
Промежуточная ( плащевая, мантийная ) зона состоит из клеток, переместившихся из вентрикулярной и субвентрикулярной зон — н ейробластов и глиобластов . Нейробласты утрачивают способность к делению и в дальнейшем дифференцируются в нейроны. Глиобласты продолжают делиться и дают начало астроцитам и олигодендроцитам . Способность к делению не утрачивают полностью и зрелые астроциты , и олигодендроциты . Новообразование нейронов прекращается в раннем постнатальном периоде. Поскольку число нейронов в головном мозге составляет примерно 1 триллион, очевидно, в среднем в течение всего пренатального периода в 1 мин формируется 2 500 000 нейронов. Из клеток плащевого слоя образуются серое вещество спинного и часть серого вещества головного мозга.
Маргинальная зона ( краевая вуаль ) формируется из врастающих в нее аксонов нейробластов и макроглии и дает начало белому веществу. В некоторых областях головного мозга клетки плащевого слоя мигрируют дальше, образуя кортикальные пластинки — скопления клеток, из которых формируется кора большого мозга и мозжечка.
По мере дифференцировки нейробласта изменяется субмикроскопическое строение его ядра и цитоплазмы. В ядре возникают участки различной электронной плотности в виде мелких зерен и нитей. В цитоплазме выявляются в большом количестве канальцы и цистерны гранулярной эндоплазматической сети, уменьшается количество свободных рибосом и полисом, значительного развития достигает аппарат Гольджи . Специфическим признаком начавшейся специализации нервных клеток следует считать появление в их цитоплазме тонких фибрилл — пучков нейрофиламентов и микротрубочек. Количество нейрофиламентов , содержащих белок — нейрофиламентный триплет, в процессе специализации увеличивается. Тело нейробласта постепенно приобретает грушевидную форму, а от его заостренного конца начинает развиваться отросток — аксон (нейрит). Позднее дифференцируются другие отростки — дендриты. Нейробласты превращаются в зрелые нервные клетки — нейроны. Между нейронами устанавливаются контакты (синапсы).
В процессе дифференцировки нейронов из нейробластов различают до- медиаторный и медиаторный периоды. Для домедиаторного периода характерно постепенное развитие в теле нейробласта органелл синтеза — свободных рибосом, а затем эндоплазматической сети. В медиатор-ном периоде у юных нейронов появляются первые пузырьки, содержащие медиатор, а в дифференцирующихся и зрелых нейронах отмечаются значительное развитие органелл синтеза и секреции (гранулярная эндоплазматическая сеть, аппарат Гольджи ), накопление медиаторов и поступление их в аксон, образование синапсов. Несмотря на то, что формирование нервной системы завершается в первые годы постнатального развития, известная пластичность центральной нервной системы сохраняется до старости. Эта пластичность может выражаться в появлении новых терминален и новых синаптических связей. Нейроны центральной нервной системы млекопитающих способны формировать новые ветви ( аксональное почкование ) и новые синапсы ( синаптическое замещение ). Пластичность проявляется в наибольшей степени в первые годы после рождения, но частично сохраняется и у взрослых — при изменении уровней гормонов, обучении новым навыкам, травме и других воздействиях. Хотя нейроны постоянны, их синаптические связи могут модифицироваться в течение всей жизни, что может выражаться, в частности, в увеличении или уменьшении их числа. Пластичность при малых повреждениях мозга проявляется в частичном восстановлении функций.
В популяции нейронов, начиная с ранних стадий развития нервной системы и в течение всего онтогенеза, имеет место массовая гибель клеток, достигающая 25-75 % всей популяции. Эта запрограммированная физиологическая гибель клеток ( апоптоз ) наблюдается как в центральной, так и в периферической нервной системе; при этом мозг теряет около 0,1 % нейронов. У человека ежегодно погибает около 10 млн нервных клеток.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом