
Нейтрофилы мигрируют из крови в инфицированные ткани благодаря хемоаттрактантам
Бактерицидный эффект нейтрофилов. Через 30—60 с после контакта мембраны нейтрофила с бактерией в нем возникает «вспышка метаболизма» с образованием перекиси водорода и суперокисных ионов, которые поражают поглощенные нейтрофилом бактерии, окисляя галогены (Сl, I) мембраны бактерий. Одновременно нейтрофилы секретируют лизоцим, лактоферрин, катионные белки, кислые и нейтральные гидролазы, поражающие фагоцитированные бактерии.
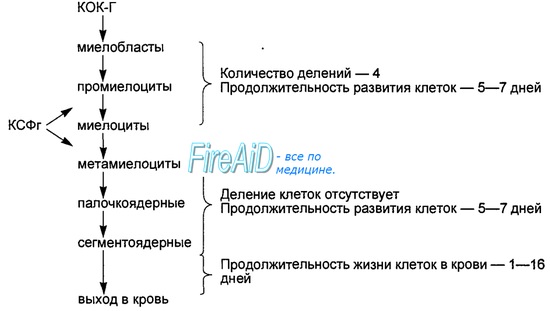
Рис. 7.5. Развитие нейтрофильных гранулоцитов. КОК-Г— колониеобразующая единица гранулоцитарная; КСФг — колониестимулируюший фактор гранулоцитарный. Выделены отделы способных к делению (КОК-Г-миелоциты) и не-размножающихся клеток (метамиелоциты—сегментоядерные) нейтрофильной линии лейкоцитов. КСФг активирует деление и усиливает функциональную активность нейтрофилов.
Нейтрофильный гранулопоэз в костном мозге представлен одновременно пролиферирующими и созревающими клетками — от миелобластов до миелоцитов включительно, и только созревающими (неделящимися) клетками — от метамиелоцитов до сегментоядерных нейтрофилов (рис. 7.5). Зрелые сегментоядерные нейтрофилы поступают из костного мозга в кровь и составляют в ней до 50—70 % всех лейкоцитов.
В небольшом количестве (1—5 %) в кровь поступают и палочкоядерные нейтрофилы. Их увеличение в крови — важный признак нарастания интенсивности нейтрофильного гранулопоэза, а также остроты воспалительного процесса. Под гранулоцитозом понимают увеличение количества нейтрофильных лейкоцитов в крови выше 10* 109/л, под нейтропенией — снижение их числа ниже 1,5 • 109/л крови.
В костном мозге в 20—25 раз больше зрелых сегментоядерных нейтрофилов, чем в крови. Это костномозговой резерв нейтрофилов. У взрослого человека он составляет 7,7 ± 1,2 • 109/кг массы тела. Гранулоцитарный колониестимулирующий фактор (КСФ-Г), бактериальный эндотоксин, N-формилпептиды бактериального происхождения, лейкотриены, катехол-амины вызывают выброс из костного мозга резервных гранулоцитов в кровь.

В крови часть гранулоцитов циркулирует, а часть — оседает у сосудистой стенки малых вен и капилляров, образуя пристеночный нециркулирующий резерв. Выброс адреналина и КСФ-Г в кровь возвращает пристеночные гранулоциты в циркулирующую кровь. За адгезию нейтрофилов и моноцитов к поверхности эндотелиальных клеток ответственны поверхностные рецепторы этих лейкоцитов — интегрины СД18, СД11а, СД11в, СД11с, а также синтезируемые в эндотелиоцитах селектины СД62 (селек-тин Р) и СД62Е (селектин Е). Последние представляют собой молекулы адгезии на поверхности эндотелиальных клеток, с которыми связываются интегрины нейтрофилов. При нарушении функций этих рецепторов нейтрофилы оказываются неспособными прикрепляться к эндотелию капилляра, а затем мигрировать из сосуда в инфицированную микроорганизмами ткань. Указанный дефект нейтрофилов способствует развитию тяжелых гнойных инфекций, что наблюдается, например, при врожденном заболевании — «синдроме недостаточности лейкоцитарной адгезии», связанном со слабым представительством (или отсутствием) интегрина СД18 на нейтрофилах.
Пристеночный резерв нейтрофилов равен 0,17 ± 0,08* 10*9 нейтрофилов/кг массы тела, циркулирующий — 0,22 ± 0,05 • 10*9 кг/массы. Средний полупериод жизни циркулирующих гранулоцитов составляет 6— 8 ч, а полный — не более 30 ч.
Нейтрофилы мигрируют из крови в инфицированные ткани благодаря хемоаттрактантам
Быстрый выход неитрофилов из сосудистого русла по направлению к очагу воспаления или инфицированным тканям является ключевым этапом в системе защиты организма от внедрения микроорганизмов. В легких 2/3 неитрофилов, находящихся в сосудистом русле, прилипают к эндотелию, образуя пристеночный пул неитрофилов. В последних исследованиях охарактеризованы молекулы, участвующие в адгезии неитрофилов к эндотелиальным клеткам. Молекулы адгезии (адгезины) неитрофилов представлены р,-интегринами (LFA-1, Мас-1, р150,95) и L-селектином]. Исключительно важная роль интегринов может быть проиллюстрирована повышенной частотой развития инфекции у больных с дефицитом интегринов. У этих больных наблюдается нейтропения и нарушение адгезии и хемотаксиса неитрофилов.
Читайте также: Липома мягких тканей надключичной области
Будучи иммобилизованными на каких-либо клетках, нейтрофилы переходят в состояние поляризации, с чего начинается процесс трансмиграции (проникновение неитрофилов из кровотока в ткани). В индукции трансмиграции неитрофилов участие L-селектина представляется крайне важным, так как его шеддинг обязателен для начала этого процесса. Большинство медиаторов, участвующих в адгезии неитрофилов, также вовлечены в регуляцию их миграции. Однако от концентрации этих медиаторов зависит то, на какую из двух активностей неитрофилов (адгезию или миграцию) будет оказано воздействие. Так, С5а в высоких концентрациях повышает адгезию неитрофилов и снижает их миграцию, в то время как обратная закономерность наблюдается при его концентрации менее 1 пМ. Более того, миграция нейтрофилов в ответ на С5а, связанный с субстратом, реализуется благодаря механизму, получившему название haptotaxis. С его помощью нейтрофилы проникают через эндотелиальный барьер, интерстициатьную ткань и, возможно, через эпителиальный слой. Различные структуры, встречающиеся на пути нейтрофилов, могут влиять на их миграционную способность. Эффективность различных хемоаттрактантов различна в условиях in vitro, a in vivo зависит от особенностей и состояния барьеров. Так, LTB4 лучше индуцирует миграцию нейтрофилов через эндотелий, нежели FMLP, в то время как последний, наоборот, индуцирует прежде всего миграцию через эпителиальный слой.
Основная роль в адгезии нейтрофилов к эпителиальным клеткам отводится экспрессированным на их мембране молекулам ICAM-1. Показано, что эта активность эпителиальных клеток повышается после их инкубации в атмосфере сигаретного дыма или после инфицирования клеток респираторными вирусами, например, вирусом парагриппа типа 2. Модуляция взаимодействия нейтрофила с эпителиальной клеткой может иметь практическое значение при респираторной инфекции, когда адгезия нейтрофилов облегчает их микробицидную активность. Наоборот, при хроническом бронхите повышенная адгезия нейтрофилов к активированным эпителиальным клеткам может индуцировать бесконтрольное развитие цитотоксичности, опосредованной нейтрофилами, и приводящей к повреждению эпителиального слоя.

Среди медиаторов воспаления следующие факторы признаны основными хемоаттрактантами для нейтрофилов, по крайней мере in vitro: С5а, NAP-l/IL-8, PAF, метаболиты арахидоновой кислоты LTB4 и 5-гидроксиэйкозатетраеновая кислота (5-НЕТЕ). TNF-a и IL-1 тоже индуцируют выход нейтрофилов из кровотока, но только in vivo. Эта активность, вероятно, связана с повышением адгезии нейтрофилов к эндотелиальным клеткам, как было обсуждено выше, и со способностью стимулировать другие воспалительные клетки к продукции хемоаттрактантов.
Альвеолярные макрофаги играют центральную роль в миграции нейтрофилов в альвеолы и в дистальные отделы дыхательных путей. Так, альвеолярные макрофаги очищают нижние отделы дыхательных путей от микроорганизмов и мелких инородных частичек. Этот процесс не требует значительной активации клеток и сопровождается лишь небольшой активацией вспомогательных клеток и иммунным ответом, постоянно наблюдаемым в легких. Однако, когда альвеолярные макрофаги поглощают микроорганизмы или какие-либо частички, они могут привлекать нейтрофилы при помощи выше перечисленных факторов: С5а, NAP-l/IL-8, PAF, LTB4, 5-НЕТЕ, TNF-a и IL-1. Наряду с этими хемоаттрактантами, альвеолярные макрофаги выделяют, по крайней мере, один низкомолекулярный ингибитор миграции нейтрофилов и их респираторного взрыва. Недавно было показано, что этот фактор представлен в секретах дыхательного тракта и его содержание находится в обратной зависимости от выраженности бронхоспазма, индуцированного ингаляцией аллергена, что подтверждает потенциальную защитную роль этого фактора при гиперреактивности бронхов. Также описаны и другие ингибиторы миграции нейтрофилов: 1) факторы сыворотки крови, угнетающие С5а-индуцированный хемотаксис нейтрофилов; 2) белок изначально выделенный из супернатантов культуры лимфоцитов, названный фактором ингибиции нейтрофилов, а в настоящее время идентифицированный как GM-CSF; 3) липоксин А4 (LXA4), метаболит арахидоновой кислоты лейкотриен А4 (LTA4).
Читайте также: Что шить из тонкой ткани
Ингибитор С5а и LXA4 обнаруживаются в бронхоальвеолярной жидкости (БАЖ), a GM-CSF может продуцироваться альвеолярными макрофагами. Когда процесс трансмиграции необходимо приостановить, ингибиторы нейтрофилов должны быстро блокировать активаторы, для того чтобы предупредить дальнейшую активацию клеток.
В связи с этим достаточно демонстративным является пример NAP-1/IL-8 — цитокина, действующего, прежде всего, на неитрофилы. Когда неитрофилы внутри сосудистого русла испытывают воздействие IL-8, на их поверхности происходит экспрессия L-селектина с последующим его шеддингом еще до контакта с эндотелиальной клеткой. В результате этого такие неитрофилы теряют способность начать трансмиграцию. Кроме того, было показано, что эритроциты с помощью высокоаффинных специфических рецепторов связывают большую часть биологически активного IL-8, содержащегося в крови. Наконец, последними исследованиями показано наличие в периферической крови антител к NAP-1/IL-8. Вместе все эти факты иллюстрируют сложность механизмов регуляции адгезии и миграции нейтрофилов.
Нейтрофилы мигрируют из крови в инфицированные ткани благодаря хемоаттрактантам
Нейтрофилы и антимикробная защита. Нейтрофил является одной из основных клеток в системе защиты легких от бактериальной и грибковой инфекции. Эксперименты на животных, также как клинические исследования, показали, что эффективность удаления бактерий из дыхательного тракта находится в прямой зависимости от чиста неитрофилов в области поражения.
Усиленное потребление клеток при тяжелых инфекциях может привести к гранулоцитопении, если продукция гранулоцитов в костном мозге по каким-то причинам подавлена. Такое явление наблюдается, например, при крупозной пневмонии у алкоголиков или лиц с дефицитом питания. Нижним лимитом считается содержание 1800 — 2000 гранулоцитов в мкл крови. При снижении количества циркулирующих гранулоцитов до 1500 клеток в мкл это проявляется нарушением течения местного острого воспалительного ответа. При падении уровня гранулоцитов в крови ниже 500 клеток в мкл прогрессивно нарастает частота инфекций.
При уровне 100 гранулоцитов в мкл все больные имеют инфекционные осложнения.

Причинами развития нейтропении могут послужить сами инфекции, в первую очередь, вирусные (HIV, HBV, EBV), а также вызванные микобактериями, грибами. Транзиторная нейтропения развивается вслед за эндотоксемией. Тяжелую нейтропению, вплоть до агранулоцитоза, может вызвать применение некоторых лечебных препаратов: сульфаниламидов, пенициллина, цефалоспоринов, фенотиазинов, антитиреоидных препаратов, хлорамфеникола.
Хорошо известно, что у больных с выраженной нейтропенией или с тяжелыми дефектами функциональной активности неитрофилов часто развивается рецидивирующая пневмония. Однако при дефектах неитрофилов инфекция не всегда развивается благодаря тому, что другие механизмы защиты выполняют компенсаторные функции. Целый ряд заболеваний связан с врожденными дефектами неитрофилов или дефицитом молекул, регулирующих их функцию. Например, дефицит CR3 — аутосомный рецессивный признак — отражается на функциях неитрофилов, связанных с их адгезией.
У таких больных нейтрофилы проявляют сниженную способность к агрегации, сниженную способность к адгезии к эндотелиальным клеткам, низкий уровень фагоцитоза опсонизирован-ных микроорганизмов, дефект распластывания, сниженный уровень спонтанной миграции и хемотаксиса. Кроме этих случаев, рецидивирующая инфекция может развиваться у больных с врожденными дефектами респираторного взрыва, например при хроническом гранулематозе и при синдроме Чедиака-Хигаси, для которого характерна дисфункция лизосом.
Нейтрофилы и повреждение легких.
В дополнение к роли нейтрофилов в инфекционных заболеваниях, эти клетки вовлечены в патогенез различных бронхолегочных заболеваний: хронического бронхита, бронхиальной астмы, муковисцидоза, респираторного дистресс синдрома у взрослых, идиопатического легочного фиброза, эмфиземы.
Протеолитические медиаторы, продуцируемые нейтрофилами для киллинга микроорганизмов, могут также действовать на окружающие ткани и, таким образом, быть вовлечены в патогенез многочисленных неинфекционных заболеваний нижних отделов респираторного тракта. Фактически отсутствуя в паренхиме легких и в дыхательных путях практически здоровых людей, нейтрофилы могут попадать в эти ткани из легочного кровотока с помощью хемоаттрактантов. Некоторые из этих факторов были обнаружены в повышенном количестве в содержимом дыхательных путей больных с заболеваниями паренхимы легких, в сочетании с повышенным количеством альвеолярных нейтрофилов.
Читайте также: Ткани для костюмов характеристика
Нейтрофилы мигрируют из крови в инфицированные ткани благодаря хемоаттрактантам
Воспалительная реакция нейтрофилов возникает, когда в крови посткапиллярных венул эти клетки «обнаруживают» небольшое количество хемокинов и других хемотаксических веществ, выделяющихся из очага инфекции. Эти растворимые эффекторы воспаления вызывают изменения состава и активности поверхностных молекул эндотелиальных клеток и нейтрофилов. Начальные взаимодействия между этими клетками (главным образом, между селектинами нейтрофилов и углеводными компонентами поверхности эндотелия) отличаются низким сродством и обратимостью.
В результате происходит так называемое перекатывание лейкоцитов по эндотелию; слабые связи образуются и распадаются, приводя к колебательному движению нейтрофилов по поверхности эндотелия. Перекатывание обеспечивает более тесный контакт нейтрофилов с такими активирующими факторами, как ФНО) или ИЛ-1, вследствие чего возникают качественные и количественные изменения Р2-интегриновых рецепторов нейтрофилов (поверхностные молекулы группы CD11/CD18).
Активированные интегриновые рецепторы прочно связывают нейтрофилы друг с другом и с эндотелиальными клетками. Окончательный результат всех этих межклеточных взаимодействий сводится к тому, что нейтрофилы распластываются на эндотелиальных клетках и образуют агрегаты друг с другом и с тромбоцитами, приводя к сужению просвета венул и снижению кровотока.
На следующем этапе интегриновые рецепторы нейтрофилов концентрируются на поверхности псевдоподий. Нейтрофилы способны перемещать свои интегриновые рецепторы по поверхности и изменять свою конфигурацию, что позволяет им проходить через контакты между эндотелиальными клетками и проникать в ткани. Затем они мигрируют в очаг инфекции по градиенту концентраций хемокинов или других хемоаттрактантов. Миграция нейтрофилов — сложный процесс, включающий циклы перемещения рецепторов, проведение сигналов в клетки и перестройку актиновых нитей цитоскелета.
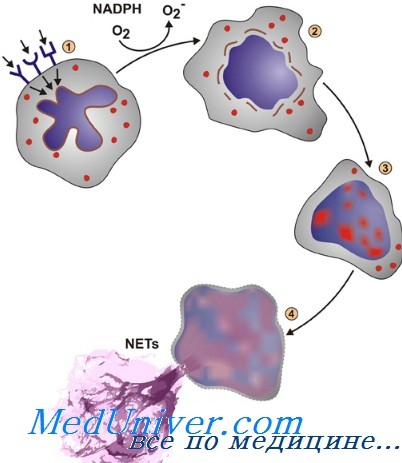
Проникновению нейтрофилов сквозь базальную мембрану сосудов и соединительную ткань способствует секреция этими клетками специфических гранул или везикулярных элементов, содержащих желатиназу, гепариназу и другие ферменты. Достигнув очага инфекции, нейтрофил с помощью Fc-участка иммуноглобулина и рецепторов комплемента, фи-бронектина и других молекул адгезии распознает патогенный агент.
Поглощаемые нейтрофилами микробы предварительно подвергаются опсонизации (подготовке к поглощению) сывороточными термостабильными и термолабильными факторами, соответственно IgG и СЗ. Опсонины облегчают фагоцитоз, при котором микроб втягивается в закрытую вакуолю, называемую фагосомой.
Образование фагосомы сопровождается двумя реакциями, необходимыми для проявления бактерицидной активности нейтрофилов, — дегрануляцией и активацией НАДФ-зависимой оксидазы. Слияние мембран гранул с мембраной фагосомы приводит к появлению в фагосоме белков, обладающих мощной антимикробной активностью. Содержимое специфических, а затем и азурофильных гранул попадает в фагосому. Одновременно в ее мембране происходит сборка и активация НАДФ-зависимой оксидазы.
Под действием этого фермента из молекулярного кислорода образуется большое количество супероксид-аниона (O2), который, в свою очередь, превращается в перекись водорода (Н2O2) и синглетный кислород. Н2O2 реагирует с O2, образуя гидроксильные радикалы. В присутствии миелопероксидазы (основного компонента азурофильных гранул) и хлоридного иона в фагосоме образуется хлорноватистая кислота (НОС1). Н2O2 и НОС1 не только обладают бактерицидной активностью, но и модулируют иммунные реакции. Эти оксиданты денатурируют белки, повышая их чувствительность к протеолизу, и, кроме того, активируют некоторые протеазы нейтрофилов.
Все это способствует разрушению патогенного агента и удалению его из очага инфекции. Оксиданты также инактивируют хемотаксические факторы, прекращая приток нейтрофилов в очаг инфекции и тем самым ослабляя процесс воспаления.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом