
С тех пор процессы пролиферации и дифференцировки клеток ушли далеко вперед, создав настоящее чудо — вас, человека. У растения жизнь начинается точно так же — с одной маленькой клетки, из которой в дальнейшем будут развиваться ткани и органы самых разных форм. Главная заслуга роста растения принадлежит образовательной ткани.
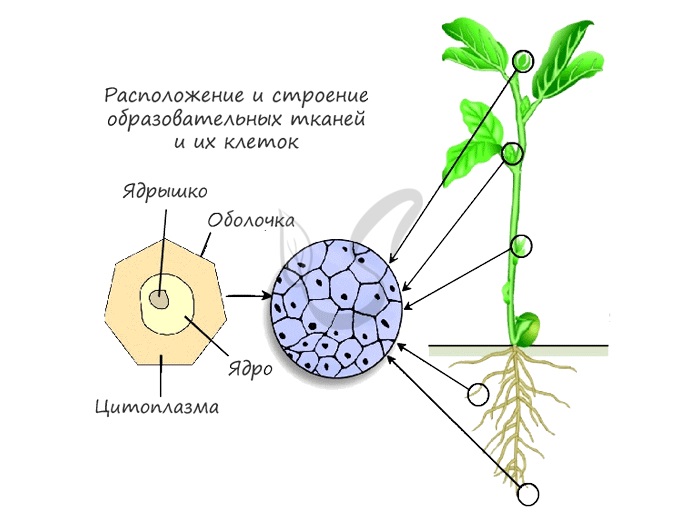
Как вы видите, на картинке схематично изображено месторасположение образовательной ткани. Главным образом это:
- Кончик побега — конус нарастания в почках
- Кончик корня — зона деления, прикрытая корневым чехликом для защиты
- Камбий — обеспечивает рост растения в ширину
- Основание междоузлий и черешков листьев — это также зоны активного роста растения
Именно в этих местах и происходит деление клеток и рост растения. Важно отметить, что сезонные изменения активности клеток камбия являются причиной возникновения годичных колец древесины. Внешний вид годичных колец обусловлен хронологической закономерностью: весной больше образуется проводящей ткани (более тонкая и рыхлая внутри), а осенью — механическая (толстая, более твердая). Именно поэтому годичные кольца на спиле дерева выглядят как чередование колец, отличающихся друг от друга.
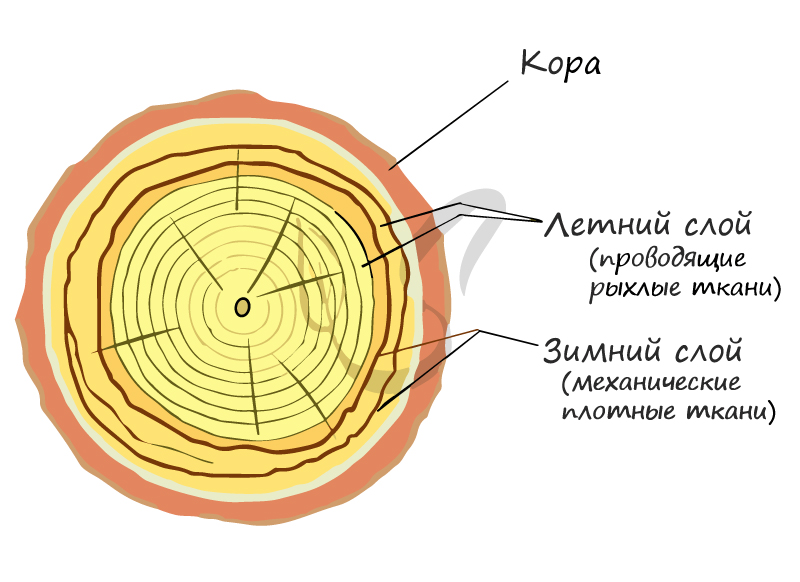
На внешний вид годичных колец оказывают весьма сильное влияние условия внешней среды. Так, при дефиците трофического компонента (питательных веществ), к примеру, у растений, растущих на болоте, годичные кольца выглядят тоньше своих обычных размеров.
Ветер также оказывает существенное влияние: при его постоянном действии происходит перераспределение древесины по стволу. Оказывая действие на крону, ветер смещает центр тяжести дерева, что сказывается на его нижележащих отделах. Они начинают компенсаторно утолщаться для предотвращения слома дерева. При постоянно дующем ветре ствол сильно искривляется, а форма кроны становится флагообразной.

Тема камбия и форм стволов растений весьма занимательна, и все-таки мы должны разобраться в строении самой образовательной ткани. Она представлена живыми мелкими быстро делящимися клетками с относительно крупным ядром. Объем цитоплазмы небольшой, она вязкая по консистенции, оболочка клетки тонкая. Это уязвимые клетки, которые растение оберегает по-своему, подобно тому, как животные оберегают только что появившееся потомство.

Другое название образовательных тканей — меристемы (с др.-греч. — «μεριστός» — делимый). По времени возникновения различают первичные и вторичные меристемы.
Первичные меристемы — закладываются в эмбриогенезе
1) Вставочные меристемы (интеркалярные) — в виде отдельных участков в зоне активного роста в разных частях растения. Такие ткани можно найти в основании междоузлий у злаков, черешков листьев у многих растений. У злаковых наблюдается быстрый рост стебля за счет множественного расположения данной ткани на стебле — «вставочный рост».
2) Прокамбий — основа будущего камбия, перицикла, окружающего проводящие ткани в один или несколько слоёв (у голосеменных). В корнях перицикл является корнеродным слоем, так как в корне с него начинается формирование осевого цилиндра, наружным слоем которого он является. В нём закладываются придаточные и боковые корни, что имеет принципиальное значение для формирования корневой системы растения.
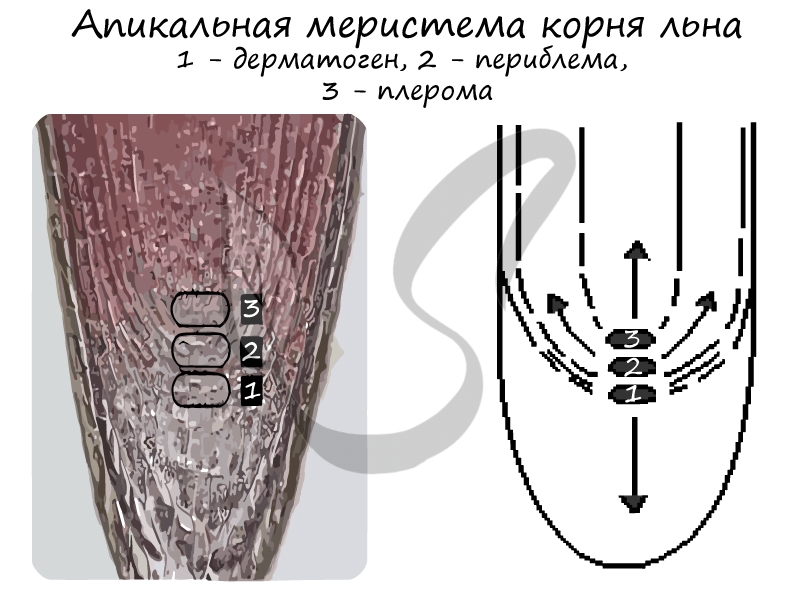
3) Верхушечные (апикальные) — формируются на верхушках стеблей и кончиках корней. В периферической части корня различают три слоя:
- Дерматоген — в дальнейшем преобразующийся в первичную покровно-всасывающую ризодерму (эпиблему или ризодерму)
- Периблема — образующая ткани первичной коры
- Плерома — внутренний слой ткани центрального осевого цилиндра
Вторичные меристемы — закладываются в постэмбриональном развитии
Камбий и феллоген (пробковый камбий) — занимают боковое положение по отношению к оси органа, обеспечивают рост вширь. Растения часто повреждаются, их задевают животные, нарушая целостность тканей и органов. На этот случай в группе вторичных меристем есть раневые меристемы, дающие начало защитной ткани в местах повреждения растения.

Топографическая классификация меристем
Спешу заверить, это отнюдь не сложная классификация, которой нужно бояться. Речь пойдет о взгляде на те же образовательные ткани с другой стороны. В переводе с греч. τόπος — место. Мы рассмотрим меристемы в соответствии с их месторасположением на растении.
- Верхушечная или апикальная (лат. apex — вершина) — расположена на кончике корня и конусе нарастания побега
- Боковая или латеральная (лат. latus — бок): камбий – обеспечивает рост стебля и корня в толщину
- Краевая или маргинальная (лат. margo — край) меристема даёт начало листовой пластинке
- Вставочная или интеркалярная (лат. inter — между и calaris — вставочный, добавочный) — расположена преимущественно у основания стеблевых междоузлий между зонами дифференцированных тканей.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Читайте также: Как называется танец тканью
Камбий — разновидность образовательных тканей растений. Характеристика и функции камбия

Все растительные ткани подразделяют на две большие группы: образовательные (или меристематические) и постоянные. Последние представляют собой уже дифференцированные группы клеток, выполняющие определенные функции (проводящую, механическую и т. д.). Постоянные ткани составляют основу растения, однако все они формируются из меристематических клеток, без которых ни эмбриональное развитие, ни рост невозможны. Одной из таких меристем является камбий.
Общая характеристика камбия
Так как главная функция камбия — это формирование проводящих растительных тканей, его наряду с прокамбием относят к васкулярным меристемам (от лат. vascularis — сосудистый). Камбий имеет вторичное происхождение и присутствует в осевых органах растений (стебли, корни). По месту локализации эту ткань относят к латеральным (боковым) меристемам.
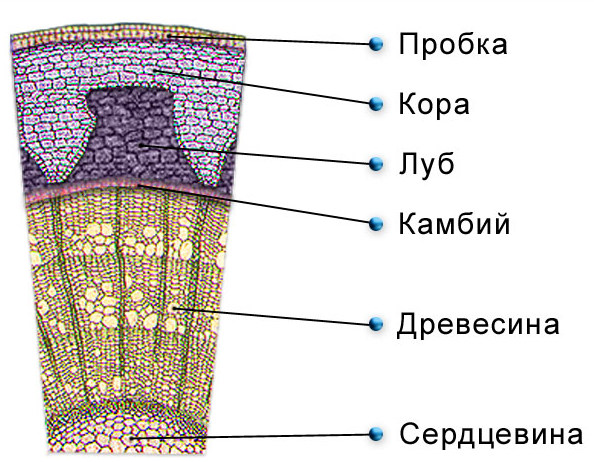
Клетки камбия живые, сильно вакуолизированные и тонкостенные. Они обладают способностью активно делиться, не истрачивая слой меристемы, в отличие от прокамбия, который со временем полностью дифференцируется в сосудистую ткань. Они имеют прозенхимную форму с заостренными концами и вытянуты вдоль поверхности осевого органа. Плоские и широкие стороны клеток обращены к проводящим тканям, а остальные стенки соприкасаются друг с другом.
При делении камбия образуются 2 типа клеток:
- инициальные — сохраняют способность к делению, полностью идентичны материнским камбиальным клеткам;
- дифференцирующиеся — способны делиться ограниченное количество (обычно 2-3) с формированием производных в виде клеток.
Инициальный слой камбия сохраняется на протяжении всей жизни растения, служа источником для образования проводящей системы. Меристема формируется непосредственно из прокамбия либо из вторичных постоянных тканей.
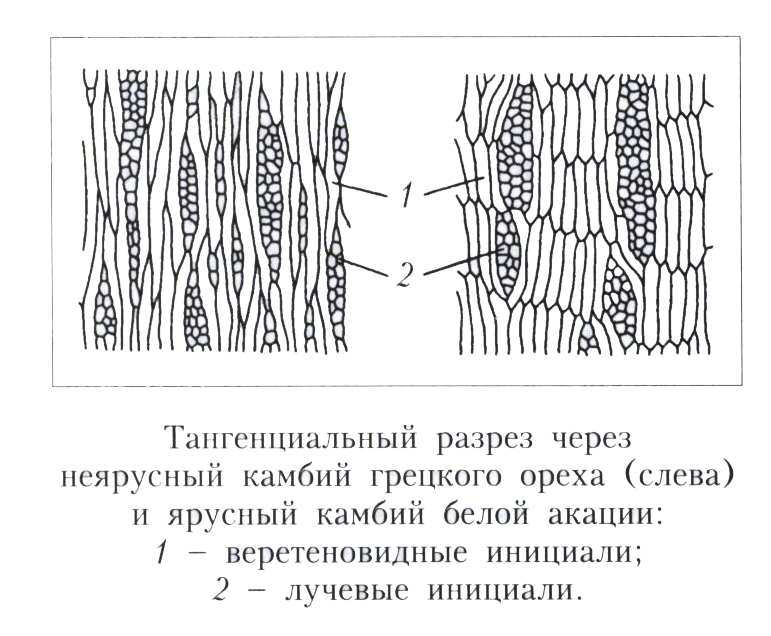
В составе камбия имеются не только веретеновидные, но и короткие лучевые клетки, дающие начало радиальным проводящим элементам — сердцевидным лучам, которые иначе называются лубодревесинными. Они обеспечивают движение веществ в горизонтальном направлении.
Функции камбия
В отличие от своего предшественника (прокамбия), камбий дает начало вторичным проводящим тканям (ксилеме и флоэме) и потому характерен преимущественно для голосеменных и двудольных покрытосеменных (как травянистых, так и древесных форм). У однодольных присутствуют только первичные проводящие ткани (метаксилема и метафлоэма), которые образуются из прокамбия. Увеличение диаметра стебля растения за счет формирование ксилемы и флоэмы из камбия называют вторичным утолщением. Таким образом меристема способствует боковому росту осевого органа.
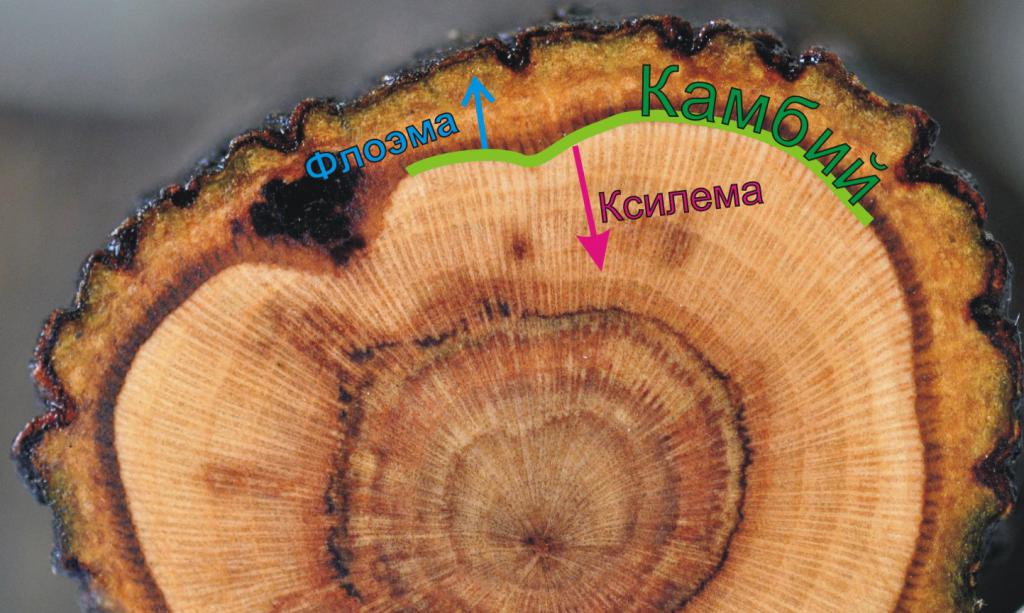
В итоге можно выделить 4 основных функции камбия:
- формирование вторичной проводящей системы;
- увеличение количества сосудистых структур по мере развития растения;
- вторичное утолщение стебля;
- увеличение диаметра осевых органов.
У растений с отмирающими побегами камбий функционирует в течение одного года, а у многолетних — на протяжении всей жизни. Прирост стебля в толщину имеет сезонный характер, так как в холодные месяцы деление клеток меристемы замирает. В стволах деревьев это приводит к формированию так называемых годичных колец.
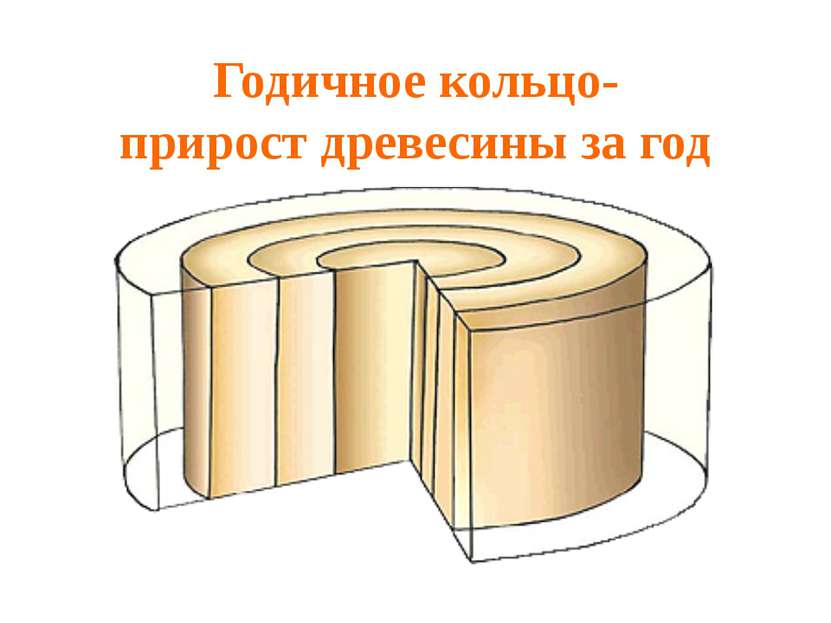
Функции камбия в стеблях травянистых и древесных растений отличаются только в количестве и характере производных. В обоих случаях работа инициального слоя приводит к откладыванию флоэмы (луба) к периферии, а ксилемы — внутрь. Разница заключается в том, что в древесном стволе элементы сосудистых тканей более жесткие, мощные и расположены компактней, чем у травянистых растений.
Как происходит вторичное утолщение?
Вторичному утолщению предшествует образование камбия, которое может происходить по трем сценариям:
- Формирование непрерывного камбиального кольца из сплошного прокамбия, которое производит непрерывные слои сосудистых тканей.
- Появление пучкового (образуется из прокамбия внутри первичных проводящих пучков) и межпучкового видов камбия, которые соединяются в сплошное кольцо и начинают откладывать ксилему и флоэму.
- Закладка меристемы происходит аналогично второму типу, но межпучковый камбий не формирует проводящие ткани, в результате чего рост ксилемы и флоэмы ограничен зоной пучков.
В последнем случае функция камбия, образующего перемычки между пучками, заключается в образовании механических элементов или паренхимы, а вторичные проводящие ткани не образуют сплошного кольца. Такая конструкция характерна для недолговечных растений с ограниченным утолщением.
Камбий в стволе древесного растения
Основной функцией камбия в дереве является формирование вторичных проводящих тканей ствола — древесины и луба. Меристема закладывается еще до окончания дифференциации первичной сосудистой системы. Возникший из ткани-предшественника камбий образует инициальный слой, дающий начало материнским клеткам древесины и луба, расположенным в пределах камбиальной зоны. Последние разрастаются в радиальном направлении, дифференцируясь в элементы сосудистой системы.
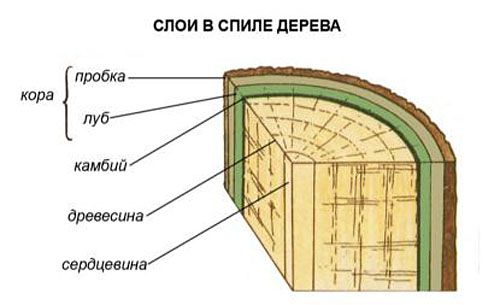
Так как клетки ксилемы и флоэмы древесных растений со временем отмирают, функция камбия заключается в поддержании проводящей системы ствола. Рабочая сосудистая зона находится вблизи васкулярной меристемы, а старые слои древесины и луба выполняют опорно-механическую и защитную роль.
Читайте также: Игрушка тигр тильда из ткани с выкройкой
Образовательные ткани — меристемы
Каждая клетка в своем развитии проходит 3 стадии: эмбриональную, стадию роста, или растяжения, и стадию дифференциации, под которой понимают совокупность изменений клеточных структур, приводящих к приобретению клеткой определенной функции, — ее специализации.
Клетки растений обладают тотипотентностью , или генетической идентичностью, так как все они являются производными зиготы, если принадлежат спорофиту, либо споры, если они составляют гаметофит. Несмотря на генетическую однородность, каждая клетка имеет специфические особенности развития в связи с тем, что гены по-разному реализуются под воздействием биохимических, в том числе гормональных и других факторов, ограничивающих развитие клетки в соответствии с ее положением в теле растения.
Рост, дифференциация и специализация составляют основу морфогенеза растений. Однако специализация, определяющая функциональную активность клетки, как мы уже говорили, обычно сопровождается утратой ее способности к делению. Одним из факторов, препятствующих делению, служит твердая оболочка клеток. У животных клетки специализированных тканей, ограниченные эластичной плазматической мембраной, многократно делятся, осуществляя так называемый диффузный рост (не делятся только клетки нервной ткани). У растений диффузный рост распространен мало. Он происходит, например, при развитии листовой пластинки. Процессы роста сосредоточены в определенных зонах тела растения, где находятся меристемы, состоящие из очень тонкостенных клеток, долго сохраняющих способность к делению. Меристемы конуса нарастания побега и кончика корня осуществляют верхушечный, или апикальный, рост, то есть надстройку новых частей растения над более старыми по времени образования, клетки которых уже приступили к дифференциации или закончили ее.
Клетка меристемы характеризуется следующими особенностями (рис. 3). Она имеет крупное ядро, занимающее около половины ее объема, в ядерной оболочке много пор, ее наружная мембрана участвует в образовании эндоплазматической сети. В гиалоплазме много диффузно расположенных рибосом. Клетка имеет пропластиды с немногочисленными тилакоидами стромы, митохондрии и диктиосомы. Вакуоли мелкие и их немного. Плазмалемма хорошо выражена. Соседние клетки соединены плазмодесмами. В клетках корневой меристемы лука их 6-7 на 1 мкм2 , у мужского папоротника — 10-20. Плазмодесмы расположены более или менее диффузно. Такое строение свойственно клеткам эвмеристемы (от греч. ей — хорошо, полностью), состоящей из изодиаметрических, в очертании многоугольных клеток. Эвмеристема участвует в образовании практически всех тканей растения за исключением проводящих.
Меристемы, образующие проводящие ткани, — прокамбий и камбий — состоят из клеток прозенхимной формы. В поперечном сечении клетки прокамбия многоугольные, клетки камбия — более или менее прямоугольные, иногда почти квадратные. И те, и другие имеют крупные вакуоли.
В связи с тем, что существенных цитологических различий между меристемами и образованными ими постоянными тканями нет, возможно новообразование меристем в теле растения. Оно происходит вследствие процесса дедифференциации живых клеток постоянных тканей, возвращающей их в состояние, близкое к эмбриональному. Эти клетки снова приобретают способность к делению. Так возникают раневые меристемы , с помощью которых восстанавливается строение поврежденных участков тела растения. На возможности новообразования меристем основано вегетативное размножение растений.
Клеткам меристем свойственно дифференцированное , или неравное деление . Клетка делится митотически на 2: одна из них остается истинной клеткой меристемы, а другая, поделившись один или несколько раз, образует клетки, вскоре приступающие к дифференциации. Неравное деление объясняет непрерывность существования меристем в растении, так как в любое время в определенных его зонах имеются клетки, способные к делению, хотя продолжительность жизни каждой из них ограничена. В этих зонах происходит постоянное взаимозамещение клеток и самовосстановление меристемы.
Меристематические клетки могут делиться в разных плоскостях по отношению к плоскости поверхности ткани или органа. Различают:
На рисунке изображены три типа деления по отношению к оси органа :
периклинальное, радиальное антиклинальное, поперечное антиклинальное
Не все клетки меристемы обладают одинаковой митотической активностью. В связи с этим в ней выделяют инициальные клетки и их производные , от которых инициальные клетки могут отличаться формой, более крупными размерами, степенью вакуолизации.
Увеличение общего объема меристемы может быть следствием активных делений либо только инициальных клеток, либо — преимущественно их производных. Эти возможности демонстрирует рис. 16. В первом случае все клетки одного вертикального ряда возникают в результате деления в горизонтальной плоскости единственной куполообразной верхушечной инициальной клетки (так нарастают, например, талломы некоторых бурых водорослей, побеги некоторых листостебельных мхов). Во втором случае инициальная клетка, отделив от себя новую клетку, на некоторое время прекращает деление. В этом случае — объем меристемы увеличивается благодаря делению ее производных. Затем инициальная клетка снова делится, отчленяя вторую клетку, которая ведет себя так же, как и первая. Такая периодичность деления инициальной клетки свойственна меристемам конусов нарастания большинства высших споровых растений. У семенных растений инициальные клетки морфологически обычно не отличимы от их производных.
Функциональная и структурная неоднородность меристем позволяет использовать для их классификации разные признаки. Морфологическая классификация основана на ориентации перегородок в делящихся клетках. С этим связаны и очертания клеток, и особенности их топографии. Выделяют следующие типы меристем.
Читайте также: Что такое lycra в составе ткани
1. Пластинчатые меристемы , состоящие из одного слоя клеток, деление которых происходит перпендикулярно поверхности органа (антиклинально). На поперечных срезах клетки обычно имеют таблитчатые очертания. Эта меристема участвует в образовании эпидермы.
2. Колончатые , или стержневые , меристемы , кубические или призматические клетки которых делятся в плоскости, перпендикулярной продольной оси органа. Клетки располагаются продольными рядами. Такая меристема образует у некоторых растений сердцевину стебля.
3. Массивные меристемы , клетки которых делятся в разных направлениях, осуществляя более или менее равномерное увеличение объема этой ткани. Очертания клеток обычно многоугольные. Из массивных меристем развивается, например, заполняющая спорангий спорогенная ткань.
Локализация в теле растения любой ткани, в том числе меристемы, подчинена принципу рационализации, способствующему наиболее эффективному функционированию тканей. Функциональные различия между меристемами определяют и их топографические особенности.
Из меристемы, точнее, из эвмеристемы, состоит развивающийся зародыш любого растения. По мере морфологической дифференциации зародыша, когда у него образуются зачатки вегетативных органов, эвмеристема сохраняется только в двух участках: на верхушке зачаточного побега, в его будущем конусе нарастания, и на кончике корня. Эти меристемы, осуществляющие рост побегов и корней в длину, называют верхушечными , или апикальными (от лат. apex — вершина).
Их непосредственные производные дифференцируются в специализированные меристемы — протодерму , основную меристему и прокамбий . Их клетки в отличие от клеток эвмеристемы более вакуолизированы, способность к делению у них ограничена. Эти меристемы часто называют полумеристемами , а также детерминированными меристемами, так как из них развиваются определенные ткани: из протодсрмы — эпидерма, из основной меристемы — механические ткани и все типы функционально разной паренхимы, из прокамбия — проводящие ткани. Топографически и онтогенетически с апикальной меристемой побега связано образование краевой ( маргинальной ) меристемы, из которой развивается пластинка листа. По мере развития она дифференцируется на те же 3 специализированные меристемы.
Наряду с апикальными меристемами многие растения имеют и боковые , или латеральные меристемы , расположенные внутри осевых органов. Они осуществляют их утолщение, наиболее сильное — у древесных форм. К латеральным меристемам относят камбий и пробковый камбий, или феллоген.
Камбий вместе с прокамбием составляет васкулярные меристемы растения (от лат. vascularis — сосудистый), участвующие в образовании проводящих тканей. Онтогенетическая связь между этими меристемами определяется тем, что в типичных случаях клетки камбия вычленяются из клеток прокамбия, при их делении периклинальными перегородками. Однако характер образования ими проводящих тканей различен: клетки прокамбия непосредственно дифференцируются в клетки первичных проводящих тканей; клетки камбия делятся перегородками, параллельными поверхности органа (периклинально), их производные дифференцируются в клетки вторичных проводящих тканей: из клеток, отложенных камбием наружу, развиваются клетки флоэмы, а из клеток, отложенных внутрь, — клетки ксилемы. Нередко камбий возникает не из прокамбия, а из постоянных тканей, живые клетки которых в этих случаях дедифференцируются. Такой камбий называют добавочным . По строению и характеру деятельности он не отличается от камбия, происхождение которого связано с прокамбием. В образовании феллогена , или пробкового камбия, также участвуют постоянные ткани. Делясь периклинально, клетки феллогена отделяют наружу будущие клетки пробки, или феллемы, а внутрь — клетки феллодермы.
Согласно генетической классификации, меристемы делят на первичные, так или иначе связанные с меристемами зародыша и апикальными меристемами, и вторичные, развивающиеся позднее, когда верхушечный рост осевых органов закончен и начинается их утолщение. Это разделение условно. Камбий, например, закладывается еще до окончания верхушечного роста, а иногда и задолго до его начала, так как имеются сведения о заложении камбия в осевых частях зимующих почек.
В табл. 2 меристемы разделены на 2 большие группы: общие и специальные. Специальные меристемы участвуют в образовании и первичных, и вторичных постоянных тканей, последние — производные камбия, независимо от его происхождения, и феллогена. При этом эпитеты «первичный» и «вторичный» несут в себе единственную информацию — последовательность образования постоянных тканей в процессе развития органов растения.
Принято выделять как особый тип интеркалярные меристемы , осуществляющие вставочный рост. Так удлиняются междоузлия на ранних стадиях развития побега, развиваются черешки листьев. Их принципиальное отличие от всех других меристем состоит в том, что наряду с недифференцированными клетками они имеют и клетки, находящиеся на разных стадиях дифференциации. Поэтому более целесообразно говорить не об интеркалярных меристемах, а о зонах интеркалярного, или вставочного роста.
Рассмотренные в этом разделе общие закономерности строения и деятельности меристем не исключают наличие у них ряда частных особенностей, присущих не только разным органам, но и разным в систематическом отношении группам растений.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом