
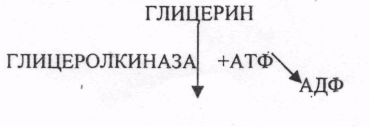
На первом этапе этих превращений затрачивается энергия: глицерин вступает в реакцию с АТФ с образованием глицерофосфата и АДФ. На следующем этапе превращений происходит окисление фосфоглицерина и его превращение в фосфодиоксиацетон (рис. 42).
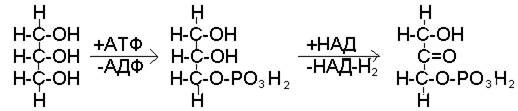
глицерин фосфоглицерин фосфодиоксиацетон
Рис. 42. Начальные этапы использования глицерина в качестве источника энергии
Перенос водорода с НАД-Н2, образующегося в ходе этой реакции, на кислород по цепи дыхательных ферментов сопряжен с ресинтезом трех молекул АТФ. Таким образом, не только компенсируются первоначальные затраты энергии, но и образуется две дополнительных молекулы АТФ.
Фосфодиоксиацетон преобразуется в свой изомер – фосфоглицериновый альдегид, дальнейшие превращения которого тождественны превращениям, происходящим в процессе окисления углеводов (рис. 43).
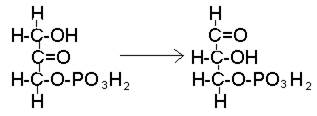
Рис. 43. Превращение фосфодиоксиацетона в фосфоглицериновый альдегид
Через несколько этапов он превращается в пировиноградную кислоту (ПВК), затем в ацетил-КоА, превращения которого завершаются в цикле трикарбоновых кислот (ЦТКК). Конечными продуктами окисления глицерина являются Н2О и СО2.
Глицерин является достаточно энергоемким веществом. При полном (до СО2и Н2О) окислении одной молекулы глицерина освобождается энергия, за счет которой может быть ресинтезирована 22 молекулы АТФ (с учетом затрат одной молекулы АТФ на начальном этапе превращений).
Дата добавления: 2015-10-29 ; просмотров: 13149 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
ОКИСЛЕНИЕ ГЛИЦЕРИНА
(тесно связанно с гликолизом)
| СН2-ОН | СН2-ОН | |||
| Глицеролфосфаткиназа | │ | α-глицеролфосфатДГ | │ | |
| ГЛЦ |   |
СН2-ОН |   |
С=О |
| АТФ АДФ | │ | НАД НАДН2 | │ | |
| СН2-ОН | СН2-О-Ф | |||
| α-глицеролфосфат | дигидроксиацетон-фосфат (ДАФ) |
ДАФ далее подвергается анаэробному и аэробному гликолизу, в последнем случае включается в ЦТК, распадается до СО2 и Н2О или при потребности организма в углеводах, посредством реакций глюконеогенеза, пореобразуется в глюкозу.
β-окисление жирных кислот
происходит в печени. Впервые изучено в 1904 году Кнопом. В 1948-1949 гг. Кеннеди и Ленинджер установили, что β-окисление жирных кислот происходит в митохондриях. В 50-х годах Линеен описал ферменты окисления жирных кислот, поэтому процесс β-окисление жирных кислот называется циклом Кноопа-Линена.
Предварительно жирные кислоты активируются в цитоплазме клетки (эндергонический процесс) т.е. процесс протекающий с повышением свободной энергии системы, требующий для своего осуществления приток энергии извне.
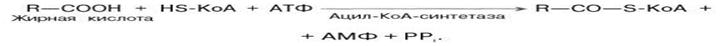
В результате образуется R-ацил-КоА, являющийся активной формой жирной кислоты.
Мембрана митохондрий не проницаема для коэнзимная формы жирной кислоты, ее транспорт в митохондрии осуществляется с помощью карнитина.
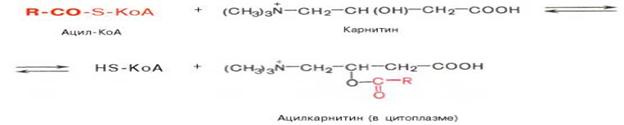
Реакция протекает при участии специфического цитоплазматического фермента карнитинацил-КоА-трансферазы. После прохождения ацилкарнитина через мембрану митохондрий происходит обратная реакция – расщепление ацилкарнитина при участии HS-KoA и митохондриальной карнитинацил-КоА-трансферазы.
Карнитин возвращается в цитоплазму клетки, а ацил-КоА подвергаются окислению в митохондриях
| О |   |
  |
О |
| ║ | ║ | ||
| R — | С |
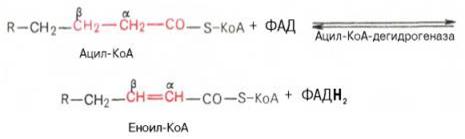

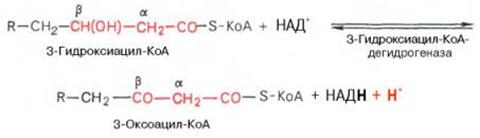
β-кетоацил КоА
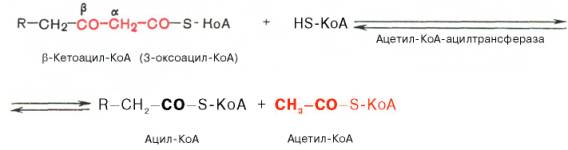
Происходит расщепление β-кетоацил КоАна ацил-КоА и ацетил-КоА, укоротившийся на два углеродных атома.
Далее цикл повторяется до тех пор, пока не образуется бутирил-КоА (4-углеродное соединение), который в свою очередь окисляется до 2 молекул ацетил-КоА.
Ацетил-КоА → в ЦТК
НАДН2,ФАДН2 → в дыхательную цепь
Энергетический баланс β-окисления
(четное число атомов углерода)
При окислении жирной кислоты, содержащей n углеродныхатомов, происходит n/2–1 цикл β-окисления (т.е. на один цикл меньше, чем n/2, так как при окислении бутирил-КоА происходит образование 2 молекул ацетил-КоА) и получится n/2 молекул ацетил-КоА. Следовательно, суммарное уравнение β-окисления пальмитиновой кислоты можно записать так:
Пальмитоил-КоА + 7ФАД + 7НАД+ + 7Н2O + 7HS-KoA ––> 8Ацетил-КоА + 7ФАДН2 + 7НАДН2.
При каждом цикле β-окисления образуются одна молекула ФАДН2 и одна молекула НАДН. Последние в процессе окисления в дыхательной цепи и сопряженного с ним фосфорилирования дают:
7ФАДН2 х 2 = 14
7НАДН2 х 3 = 21 35 молекул АТФ ( в дыхательную цепь)
8 ацетил-КоА х 12 = 96 молекулАТФ (в ЦТК)
ИТОГО: 35+96=131-1(на образование активной формы жк) = 130 молекул АТФ.
Для ненасыщенной жк кол-во двойных связей х 2 = количество молекул АТФ.
Окисление высших жирных кислот
Жировая ткань, состоящая из адипозоцитов, выполняет специфическую роль в липидном обмене. Около 65% массы жировой ткани приходится на долю отложенных в ней триацилглицеролов (ТАГ) — они представляют собой форму запасания энергии и выполняют в обмене жиров такую же функцию, как гликоген печени в обмене углеводов. Отложенные жиры в жировой ткани служит источником эндогенной воды и энергетическим резервом для организма человека. ТАГ используется в организме после предварительного расщепления (липолиза), в ходе которого освобождаются глицерин и свободные жирные кислоты. В клетках жировой ткани при участии липаз происходит распад ТАГ. Липаза находится в неактивной форме, она активируется гормонами (адреналином, норадреналином, глюкагоном, тироксином, глюкокортикоидами, СТГ, АКТГ) в ответ на стресс, голодание, охлаждение продуктами реакции являются моноацилглицерин и ВЖК. ВЖК с помощью альбуминов переносятся кровью к клеткам тканей, органов, где происходит их окисление.
Окисление высших жирных кислот.
-фосфолипиды клеточных биомембран
Окисление ВЖК происходят в митохондриях клеток, и называется бетта окислением. Доставка их к тканям и органам происходит при участии альбумина, а транспорт из цитоплазмы в митохондрии при участии карнитина. Процесс бета-окисления ВЖК складывается из следующих этапов:
-активация ВЖК на наружной поверхности мембраны митохондрий при участии АТФ, конзима А и ионов магния с образованием активной формы ВЖК (ацил — КоА).
-транспорт жирных кислот внутрь митохондрий возможен при присоединении активной формы жирной кислоты к карантину, находящемуся на наружной поверхности внутренней мембраны митохондрий. Образуется ацил-карнитин, обладающий способностью проходить через мембрану. На внутренней поверхности комплекс распадается и карнитин возвращается на наружную поверхность мембраны.

-внутримитохондриальное окисление жирных кислот состоит из последовательных ферментативных реакций. В результате одного завершенного цикла окисления происходит отщепление от жирой кислоты одной молекулы ацетил-КоА, т.е. укорочение жирнокислотной цепи на два углеродных атома.Приэтом в результате двух дегидрогеназных реакций восстанавливается ФАД до ФАДН2 и НАД + до НАДН2. Т.о. завершая 1 цикл бега — окисления ВЖК, в результате которого ВЖК укоротилось на 2 углеродных звена. При бета -окислении выделилось 5АТФ и 12АТФ выделилось при окислении АЦЕТИЛ-КОА в ЦТК и сопряженных с ним ферментов дыхательной цепи. Окисление ВЖК будет происходить циклически одинаково, но только до последней стадии — стадии превращения масляной кислоты (БУТИРИЛ-КОА), которая имеет свои особенности, которые необходимо учитывать при подсчёте суммарного энергетического эффекта окисления ВЖК, когда в результате одного цикла образуется 2 молекулы АЦЕТИЛ-КОА, одна из них проходила бета -окисление с выделением 5АТФ, а другая нет.
ОКИСЛЕНИЕ ВЖК, ИМЕЮЩИХ НЕЧЕТНОЕ КОЛИЧЕСТВО УГЛЕРОДНЫХ ЗВЕНЬЕВ В ЦЕПИ
Такие ВЖК поступают в организм человека в составе пищи с мясом жвачных животных, растений, морских организмов. Окисление таких ВЖК происходит также как и ВЖК, имеющих чётное количество углеродных звеньев в цепи, но только до последней стадии — стадии превращения ПРОПИОНИЛ-КОА. которая имеет свои особенности.

Т.о. образуется СУКЦИНИЛ-КОА, который в дальнейшем окисляется в МИТОХОНДРИЯХ с участием ферментов ЦТК КРЕБСА и сопряженных с ним ферментов дыхательной цепи.
Окисление глицерина
Окисление глицерина в тканях тесно связано с ГЛИКОЛИЗОМ, в который вовлекаются метаболиты обмена глицерина по следующей схеме:

При окислении глицерина образовались конечные продукты:
Окисление глицерина в тканях. Энергетический эффект этого процесса.
В результате гидролиза жира образуются общие метаболиты: глицерины и ВЖК, окисление которых сопровождается образованием конечных продуктов -воды и углекислого газа -и выделением энергии в форме АТФ. Окисление глицеринов в тканях тесно связано с ГЛИКОЛИЗОМ, в который вовлекаются метаболиты обмена глицерина по следующей схеме:
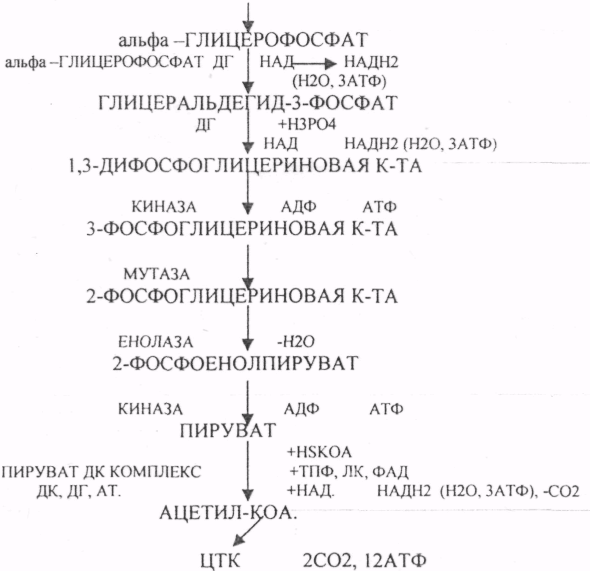
Т.о. при окислении глицерина образовались конечные продукты:
3. Альфа-КЕТОГЛУТАРАТА АТФ на этапе превращения:
3. 1,3-ДИФОСФОГЛИЦЕРИНОВОЙ К-ТЫ (СУБСТРАТНОЕФОСФОРИЛИРОВАНИЕ)
4. 2-ФОСФОЕНОЛПИРУВАТА (СУБСТРАТНОЕ ФОСФОРИЛИРОВАНИЕ)
8. СУКЦИНИЛ-КОА (СУБСТРАТНОЕ ФОСФОРИЛИРОВАНИЕ)
Биосинтез высших жирных кислот в тканях. Биосинтез жиров в печени и жировой ткани.
Биосинтез ВЖК протекает в ЭПС клеток. Исходным материалом для синтеза заменимых ВЖК является АЦЁТИЛ-КОА.
Условия для биосинтеза ВЖК:
1.Наличие АТФ, СО2, Н2О, НАДФ*Н2, поступающий из ГЕКСОЗОМОНОФОСФАТНОГО пути превращения глюкозы.
2.Наличие специальных белков-переносчиков (HS -АПБ).
3.Наличие специальных ферментов синтеза.
Процесс биосинтеза циклический. Каждый цикл включает в себя 6 этапов. АЦЕТИЛ-КОА используется на первом этапе, как «затравка» синтеза.
Первый этап. Этап образования 3-углеродного соединения — МАЛОНИЛ-КОА.
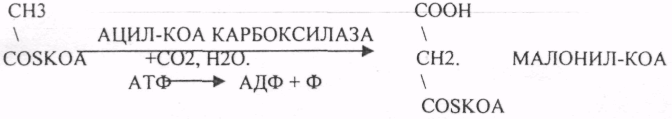
Второй этап. Этап переноса МАЛОНИЛА и АЦЕТИЛА на специальные белки — HS-АПБ.
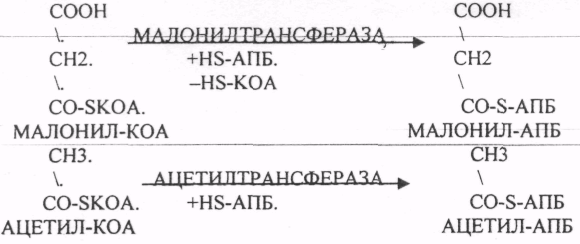
Третий этап. Этап конденсации МАЛОНИЛА-АПБ и АЦЕТИЛА-АПБ с участием ТРАНСФЕР АЗЫ — СИНТАЗЫ.
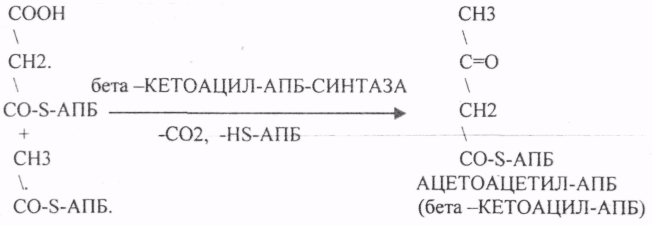
Четвёртый этап. Этап восстановления бета -КЕТОАЦИЛ-АПБ.
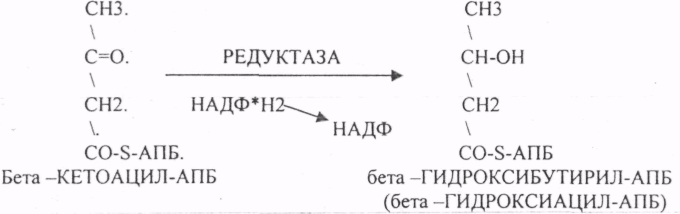
Пятый этап. Этап ДЕГИДРАТАЦИИ бета-ГИДРОКСИАЦИЛ-АПБ.

Шестой этап. Этап восстановления ЕНОИЛАЦИЛ-АПБ.

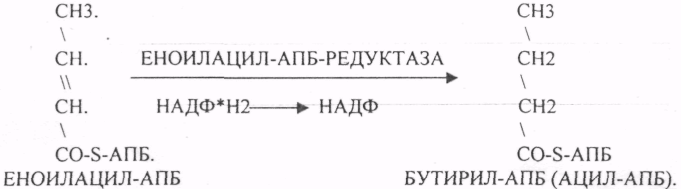 Т.о. завершается 1 цикл синтеза ВЖК образованием масляной кислоты (БУТИРИЛ-АПБ). В дальнейшем последовательно и циклично к 4-углеродному фрагменту будут присоединяться молекулы МАЛОНИЛ-КОА. Для подсчёта количества циклов синтеза ВЖК, подсчёта молекул АТФ, МАЛОНИЛ-КОА можно пользоваться формулой: N/2 — 1, где N-это число углеродных звеньев в молекуле ВЖК. Завершается биосинтез любой ВЖК в тканях ДЕАЦИЛАЗНОЙ реакцией.
Т.о. завершается 1 цикл синтеза ВЖК образованием масляной кислоты (БУТИРИЛ-АПБ). В дальнейшем последовательно и циклично к 4-углеродному фрагменту будут присоединяться молекулы МАЛОНИЛ-КОА. Для подсчёта количества циклов синтеза ВЖК, подсчёта молекул АТФ, МАЛОНИЛ-КОА можно пользоваться формулой: N/2 — 1, где N-это число углеродных звеньев в молекуле ВЖК. Завершается биосинтез любой ВЖК в тканях ДЕАЦИЛАЗНОЙ реакцией.
Так в организме человека происходит синтез всех заменимых ВЖК — всех предельных и непредельных ВЖК, имеющих одну двойную связь.
Холестерол. Его химическое строение, биосинтез и биологическая роль. Причины гиперхолестеринемии.
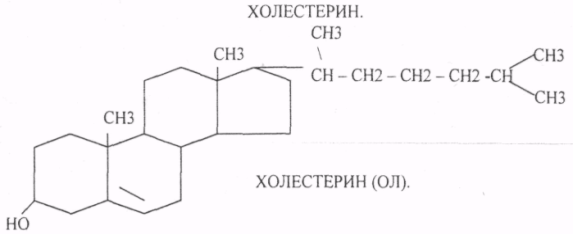
Холестерин является одноатомным циклическим спиртом, который в тканях легко образует ХОЛЕСТЕРИДЫ. В организм человека поступает в составе пищи и синтезируется г.о. в печени, тонком отделе кишечника и коже.
Биологическая роль холестерина:
1.Структурная. Свободный холестерин является, обязательным структурным компонентом мембран клеток.
2.Метаболическая. Холестерин является предшественником биологически активных веществ: витамина D3,СТЕРОИДНЫХ гормонов (АНДРОГЕНОВ, ЭСТРОГЕНОВ, КОРТИКОИДОВ) При окислении холестерина в печени при участии ЦИТОХРОМА Р-450 образуются желчные кислоты. В свободном виде холестерин транспортируется по организму с помощью транспортных ЛИПОПРОТЕИНОВ крови. Источники холестерина:
1. Пища. За сутки в организм взрослого человека поступает 0,3гр. холестерина.
2. У человека в среднем с массой 65-70кг за сутки синтезируется 3.5 -4,2гр. холестерина. Печень занимает главное место в синтезе холестерина (85%), холестерин синтезируется в кишечнике (10%) и коже (5%). Процесс биосинтеза многоступенчат:

Образовавшийся в результате распада мембранных ЛИПИДОВ, а также излишки холестерина с помощью ЛПВП доставляется для окисления в печень, и в составе желчных кислот удаляется из организма с каловыми массами. При поражении печени и кишечника нарушается образование и транспорт ЛП крови. При поражении печени и желчевыводяицих путей нарушается образование и экскреция желчных кислот, участвующих в переваривании жиров пищи. В случае нарушения оттока желчи происходит насыщение её холестерином, что ведёт в этих условиях к застою и образованию холестериновых камней. Развивается ЖЕЛЧЕКАМЕННАЯ БОЛЕЗНЬ. В крови отмечается ГИПЕРХОЛЕСТЕРИНЕМИЯ.
Основные проявления патологии липидного обмена и возможные причины их возникновения на различных этапах обмена веществ. Образование кетоновых тел в тканях. Кетоацидоз. Биологическое значение кетоновых тел.
1 .На этапе поступления жиров с пищей:
A. Обильная жирная пища на фоне ГИПОДИНАМИИ приводит к развитию АЛИМЕНТАРНОГО ОЖИРЕНИЯ.
B. Недостаточное поступление жиров или их отсутствие приводит к ГИПО- и АВИТАМИНОЗАМ A, D, Е, К. Могут развиваться ДЕРМАТИТЫ, СКЛЕРОЗ сосудов. Также нарушается процесс синтеза ПРОСТАГЛАНДИНОВ.
C. Недостаточное поступление с пищей ЛИПОТРОПНЫХ(холин, серин, инозит, витамины В12, В6) веществ приводит к развитию жировой инфильтрации тканей.
A. При поражении печени и кишечника нарушается образование и транспорт ЛП крови.
B. При поражении печени и желчевыводяицих путей нарушается образование и экскреция желчных кислот, участвующих в переваривании жиров пищи. Развивается ЖКБ. В крови отмечается ГИПЕРХОЛЕСТЕРИНЕМИЯ.
C. Если поражается слизистая оболочка кишечника и нарушается выработка и поступление ферментов ПЖЖ, содержание жира в кале увеличивается. Если содержание жира будет превышать 50%, развивается СТЕАТОРЕЯ. Кал становится бесцветным.
D. Наиболее часто в последнее время среди населения встречается поражение бета -клеток ПЖЖ, что ведет к развитию сахарного диабета, который сопровождается интенсивным окислением в клетках белков и жиров. В крови у таких больных отмечается ГИПЕРКЕТОНЕМИЯ, ГИПЕРХОЛЕСТЕРИНЕМИЯ. Кетоновые тела и холестерин синтезируются из АЦЕТИЛ-КОА.
3.На этапе обмена холестерина наиболее частым заболеванием является АТЕРОСКЛЕРОЗ. Болезнь развивается тогда, когда между клетками тканей и ЛП крови растёт содержание АТЕРОГЕННЫХ ФРАКЦИЙ и понижается содержание ЛПВП, назначение которых удалять холестерин из клеток тканей в печень для последующего его окисления. Все ЛП за исключением ХИЛОМИКРОНОВ быстро метаболизируются. ЛПНП задерживаются в сосудистой стенке. Они содержат много ТРИГЛИЦЕРИДОВ и ХОЛЕСТЕРИНА. Они, фагоцитируясь, разрушаются ферментами ЛИЗОСОМ, за исключением холестерина. Он накапливается в клетке в большом количестве. Холестерин откладывается в межклеточном пространстве и инкапсулируется соединительной тканью. В сосудах образуются АТЕРОСКЛЕРОТИЧЕСКИЕ БЛЯШКИ.
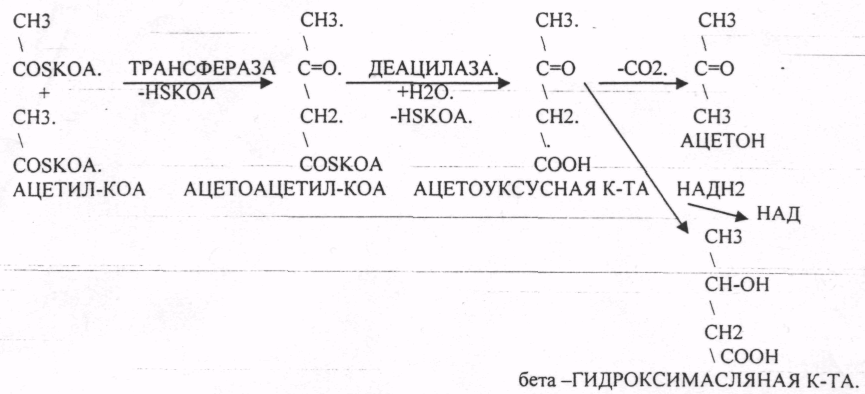
Кетоновые тела (не более 0,1 г/л) — ацетон, ацетоуксусная кислота, бета -гидроксимасляная кислота. При дефиците углеводов в клетке жиры не могут полностью окисляться, и избыток ацетил-КоА компенсируется образованием кетоновых тел. Опасно в отношении КЕТОАЦИДОЗА.
Дата добавления: 2019-07-15 ; просмотров: 2947 ; Мы поможем в написании вашей работы!