
Окислительное декарбоксилирование пировиноградной кислоты
Окисление пирувата до ацетил-КоА происходит при участии ряда ферментов и коферментов, объединенных структурно в мультиферментную систему, получившую название «пируватдегидрогеназный комплекс».
На I стадии этого процесса пируват (рис. 10.8) теряет свою карбоксильную группу в результате взаимодействия с тиаминпирофосфатом (ТПФ) в составе активного центра фермента пируватдегидрогеназы (E1). На II стадии оксиэтильная группа комплекса E1–ТПФ–СНОН–СН3 окисляется с образованием ацетильной группы, которая одновременно переносится на амид липоевой кислоты (кофермент), связанной с ферментом дигидроли-поилацетилтрансферазой (Е2). Этот фермент катализирует III стадию – перенос ацетильной группы на коэнзим КоА (HS-KoA) с образованием конечного продукта ацетил-КоА, который является высокоэнергетическим (макроэргическим) соединением.
На IV стадии регенерируется окисленная форма липоамида из восстановленного комплекса дигидролипоамид–Е2. При участии фермента дигидролипоилдегидрогеназы (Е3) осуществляется перенос атомов водорода от восстановленных сульфгидрильных групп дигидролипоамида на ФАД, который выполняет роль простетической группы данного фермента и прочно с ним связан. На V стадии восстановленный ФАДН2 дигидро-липоилдегидрогеназы передает водород на кофермент НАД с образованием НАДН + Н + .
Процесс окислительного декарбоксилирования пирувата происходит в матриксе митохондрий. В нем принимают участие (в составе сложного мультиферментного комплекса) 3 фермента (пируватдегидрогеназа, ди-гидролипоилацетилтрансфераза, дигидролипоилдегидрогеназа) и 5 кофер-ментов (ТПФ, амид липоевой кислоты, коэнзим А, ФАД и НАД), из которых три относительно прочно связаны с ферментами (ТПФ-E1, ли-поамид-Е2 и ФАД-Е3), а два – легко диссоциируют (HS-KoA и НАД).
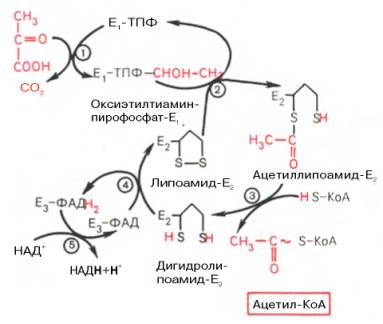
Рис. 10.8. Механизм действия пируватдегидрогеназного комплекса.
Е1 — пируватдегидрогеназа; Е2 — ди-гидролипоилацетилтрансфсраза; Е3 -дигидролипоилдегидрогеназа; цифры в кружках обозначают стадии процесса.
Все эти ферменты, имеющие субъединичное строение, и коферменты организованы в единый комплекс. Поэтому промежуточные продукты способны быстро взаимодействовать друг с другом. Показано, что составляющие комплекс полипептидные цепи субъединиц дигидролипоил-ацетилтрансферазы составляют как бы ядро комплекса, вокруг которого расположены пируватдегидрогеназа и дигидролипоилдегидрогеназа. Принято считать, что нативный ферментный комплекс образуется путем самосборки.
Суммарную реакцию, катализируемую пируватдегидрогеназным комплексом, можно представить следующим образом:
Пируват + НАД + + HS-KoA –> Ацетил-КоА + НАДН + Н + + СO2.
Реакция сопровождается значительным уменьшением стандартной свободной энергии и практически необратима.
Образовавшийся в процессе окислительного декарбоксилирования аце-тил-КоА подвергается дальнейшему окислению с образованием СО2 и Н2О. Полное окисление ацетил-КоА происходит в цикле трикарбоновых кислот (цикл Кребса). Этот процесс, так же как окислительное декарбо-ксилирование пирувата, происходит в митохондриях клеток.
Окислительное декарбоксилирование пировиноградной кислоты (пвк)
Окисление ПВК до ацетил-КоА происходит при участии ряда ферментов и коферментов, объединенных структурно в мультиферментную систему, получившую название «пируватдегидрогеназный комплекс».
Процесс окислительного декарбоксилирования ПВК происходит в матриксе митохондрий. В нем принимают участие (в составе пируватдегидрогеназного комплекса) 3 фермента (пируватдегидрогеназа, дигидролипоилдегидрогеназа и дигидролипоилацетилтрансфераза) и 5 коферментов (тиаминпирофосфат (ТПФ (витамин В1)), амид липоевой кислоты, коэнзим А (СоА (витамин В3)), ФАД (флавинадениндинуклеотид (витамин В2)),НАД + (никотинамидадениндинуклеотид (витамин РР)).
Суммарную реакцию, катализируемую пируватдегидрогеназным комплексом, можно представить следующим образом:
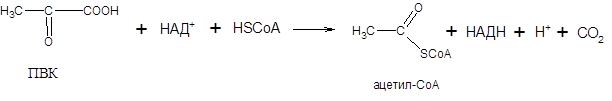
Образовавшийся в процессе окислительного декарбоксилирования ацетил-СоА окисляется далее в цикле трикарбоновых кислот с образованием СО2 и АТФ.
Читайте также: Саки ткани для мебели
42.Окислительное декарбоксилирование пировиноградной кислоты. Его биологическое значение.
Окислительное декарбоксилирование пировиноградной кислоты (пвк)
Окисление ПВК до ацетил-КоА происходит при участии ряда ферментов и коферментов, объединенных структурно в мультиферментную систему, получившую название «пируватдегидрогеназный комплекс».
Процесс окислительного декарбоксилирования ПВК происходит в матриксе митохондрий. В нем принимают участие (в составе пируватдегидрогеназного комплекса) 3 фермента (пируватдегидрогеназа, дигидролипоилдегидрогеназа и дигидролипоилацетилтрансфераза) и 5 коферментов (тиаминпирофосфат (ТПФ (витамин В1)), амид липоевой кислоты, коэнзим А (СоА (витамин В3)), ФАД (флавинадениндинуклеотид (витамин В2)),НАД + (никотинамидадениндинуклеотид (витамин РР)).
Суммарную реакцию, катализируемую пируватдегидрогеназным комплексом, можно представить следующим образом:
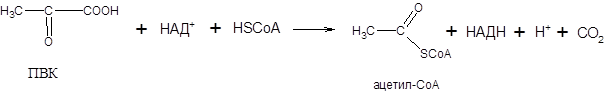
Образовавшийся в процессе окислительного декарбоксилирования ацетил-СоА окисляется далее в цикле трикарбоновых кислот с образованием СО2 и АТФ.
Биологическая роль окислительного декарбоксилирования пирувата заключается в том, что оно является важным этапом катаболизма, позволяющим включаться в цикл Кребса тем веществам, при распаде которых образуется ПВК. Образовавшаяся молекула НАДН2 окисляется в длинной дыхательной цепи с образованием 3-х молекул АТФ. Окислительное декарбоксилирование пирувата протекает внутри митохондрий.
43.Специфические и общие пути катаболизма углеводов, липидов, белков. Цикл трикарбоновых кислот (цикл Кребса), его роль в обмене углеводов, липидов, аминокислот.
В катаболизме различают три стадии:
1). Полимеры превращаются в мономеры (белки – в аминокислоты, углеводы в моносахариды, липиды – в глицерол и жирные кислоты). Химическая энергия при этом рассеивается в виде тепла.
2). Мономеры превращаются в общие продукты, в подавляющем большинстве в ацетил-КоА. Химическая энергия частично рассеивается в виде тепла, частично накапливается в виде восстановленных коферментных форм (НАДН, ФАДН2), частично запасается в макроэргических связях АТФ (субстратное фосфорилирование).
1-ая и 2-ая стадии катаболизма относятся к специфическим путям, которые уникальны для метаболизма белков, липидов и углеводов.
3). Заключительный этап катаболизма, сводится к окислению ацетил-КоА до СО2 и Н2О в реакциях цикла трикарбоновых кислот (цикла Кребса) – общий путь катаболизма. Окислительные реакции общего пути катаболизма сопряжены с цепью тканевого дыхания. При этом энергия (40-45%) запасается в виде АТФ (окислительное фосфорилирование).
В результате специфических и общих путей катаболизма биополимеры (белки, углеводы, липиды) распадаются до СО2, Н2О и NH3, которые являются основными конечными продуктами катаболизма.
ЦИКЛ КРЕБСА(цикл лимонной и трикарбоновой кислот), система биохимических реакций, посредством которой большинство организмов ЭУКАРИОТОВ получают основную энергию в результате окисления пищи. Происходит в КЛЕТКАХ МИТОХОНДРИЙ. Включает несколько химических реакций, в результате которых высвобождается энергия. Этот процесс называется системой ПЕРЕНОСА ЭЛЕКТРОНОВ по аналогии с переходом АДЕНОЗИНТРИФОСФАТА (АТФ) в АДЕНОЗИН ДИФОСФАТ (АДФ). АТФ обеспечивает реакции МЕТАБОЛИЗМА химической энергией. Цикл Кребса важная часть процесса ДЫХАНИЯ и ОБМЕНА ВЕЩЕСТВ клетки.
Цикл трикарбоновых кислот (цикл Кребса)
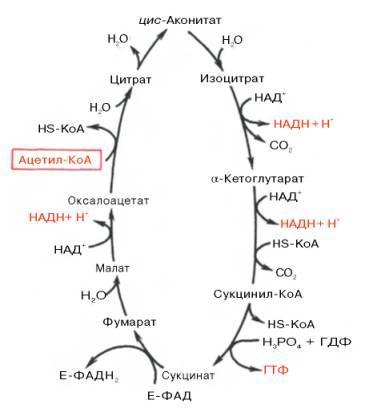
образовавшийся в результате окислительного декарбоксилирования пирувата ацетил-КоА вступает в цикл Кребса. Данный цикл состоит из восьми последовательных реакций
Читайте также: Выкройки сердечек из ткани разных размеров
В первой реакции, катализируемой ферментом цитратсинтазой, ацетил-КоА конденсируется с оксалоацетатом. В результате образуется лимонная кислота:
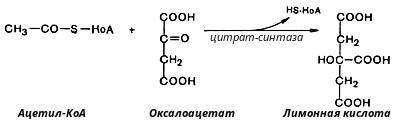
По-видимому, в данной реакции в качестве промежуточного продукта образуется связанный с ферментом цитрил-КоА. Затем последний самопроизвольно и необратимо гидролизуется с образованием цитрата и HS-KoA.
Во второй реакции цикла образовавшаяся лимонная кислота подвергается дегидратированию с образованием цис-аконитовой кислоты, которая, присоединяя молекулу воды, переходит в изолимонную кислоту. Катализирует эти обратимые реакции гидратации-дегидратации фермент аконитат-гидратаза:
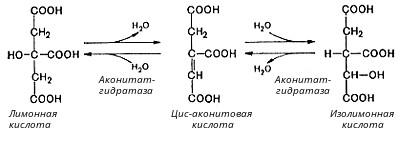
В третьей реакции, которая, по-видимому, лимитирует скорость цикла Кребса, изолимонная кислота дегидрируется в присутствии НАД-зависимой изоцитратдегидрогеназы:
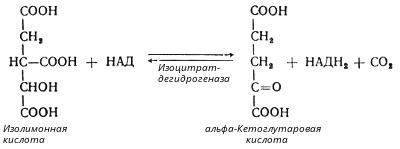
(В тканях существует два типа изоцитратдегидрогеназ: НАД- и НАДФ-зависимые. Установлено, что роль основного катализатора окисления изолимонной кислоты в цикле Кребса выполняет НАД-зависимая изоцитратдегидрогеназа.)
В ходе изоцитратдегидрогеназной реакции изолимонная кислота декарбоксилируется. НАД-зависимая изоцитратдегидрогеназа является аллостерическим ферментом, которому в качестве специфического активатора необходим АДФ. Кроме того, фермент для проявления своей активности нуждается в ионах Mg2+или Мn2+.
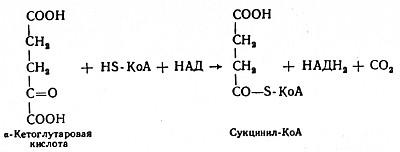
В четвертой реакции происходит окислительное декарбоксилирование α-кетоглутаровой кислоты до сукцинил-КоА. Механизм этой реакции сходен с реакцией окислительного декарбоксилирования пирувата до ацетил-КоА. α-Кетоглутаратдегидрогеназный комплекс напоминает по своей структуре пируватдегидрогеназный комплекс. Как в одном, так и в другом случае в ходе реакции принимают участие пять коферментов: TДФ, амид липоевой кислоты, HS-KoA, ФАД и НАД. Суммарно данную реакцию можно написать так:
Пятая реакция катализируется ферментом сукцинил-КоА-синтетазой. В ходе этой реакции сукцинил-КоА при участии ГДФ и неорганического фосфата превращается в янтарную кислоту (сукцинат). Одновременно происходит образование высокоэргической фосфатной связи ГТФ1 за счет высокоэргической тиоэфирной связи сукцинил-КоА:
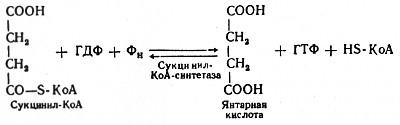
(Образовавшийся ГТФ отдает затем свою концевую фосфатную группу на АДФ, вследствие чего образуется АТФ. Образование высокоэргического нуклеозидтрифосфата в ходе сукцинил-КоА-синтетазной реакции — пример фосфорилирования на уровне субстрата.)
В шестой реакции сукцинат дегидрируется в фумаровую кислоту. Окисление сукцината катализируется сукцинатдегидрогеназой, в молекуле которой с белком ковалентно связан кофермент ФАД:
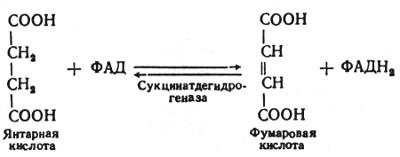
В седьмой реакции образовавшаяся фумаровая кислота гидратируется под влиянием фермента фумаратгидратазы. Продуктом данной реакции является яблочная кислота (малат). Следует отметить, что фумаратгидратаза обладает стереоспецифичностью, — в ходе данной реакции образуется L-яблочная кислота:
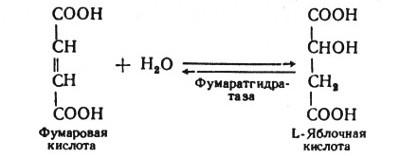
Наконец, в восьмой реакции цикла трикарбоновых кислот под влиянием митохондриальной НАД-зависимой малатдегидрогеназы происходит окисление L-малата в оксалоацетат:
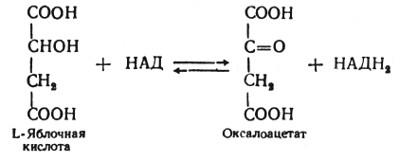
Как видно, за один оборот цикла, состоящего из восьми ферментативных реакций, происходит полное окисление («сгорание») одной молекулы ацетил-КоА. Для непрерывной работы цикла необходимо постоянное поступление в систему ацетил-КоА, а коферменты (НАД и ФАД), перешедшие в восстановленное состояние, должны снова и снова окисляться. Это окисление осуществляется в системе переносчиков электронов (или в цепи дыхательных ферментов), локализованной в митохондриях.

СВЯЗЬ МЕЖДУ ОБМЕНОМ УГЛЕВОДОВ И ЖИРОВ
Единство в обмене углеводов и жиров доказывается возникновением общих промежуточных продуктов распада. При распаде углеводов образуется пировиноградная кислота, а из нее -активная уксусная кислота -ацетил-КоА, который может быть использован в синтезе жирных кислот. Последние при своем распаде дают ацетил-КоА.