
Мышечные ткани — это ткани, для которых способность к сокращению является главным свойством. Мышечные ткани составляют активную часть опорно-двигательного аппарата (пассивной частью являются кости, соединения костей).
Общими свойствами всех мышечных тканей является сократимость и возбудимость. К данной группе тканей относятся гладкая, поперечнополосатая скелетная и поперечнополосатая сердечная мышечные ткани. Клетки мышечной ткани имеют хорошо развитый цитоскелет, содержат много митохондрий.

Гладкая (висцеральная) мускулатура
Эта мышечная ткань встречается в стенках внутренних органах (бронхи, кишечник, желудок, мочевой пузырь), в стенках сосудов, протоках желез. Эволюционно является наиболее древним видом мускулатуры.
Состоит из веретенообразных миоцитов — коротких одноядерных клеток. Между клетками имеются межклеточные контакты — нексусы (лат. nexus — связь). Благодаря нексусам возбуждение, возникшее в одной клетке, волнообразно распространяется на все остальные клетки.

Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы внутренних органов (к примеру мочевого пузыря), сокращается медленно, практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает — сокращается и утомляется быстро.
Осуществляется сокращение с помощью клеточных органоидов — миофиламентов, которые расположены в клетке хаотично и не имеют такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их тоже изучим).
Особо заметим, что в гладкой мышечной ткани миофиламенты собираются в миофибриллы только во время сокращения. У таких временных миофибрилл не может быть регулярной организации, а значит ни у таких миофибрилл, ни у гладких миоцитов не может быть поперечной исчерченности.
Гладкая мышечная ткань сокращается непроизвольно (неподвластна воле человека). Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой. К примеру невозможно по желанию сузить или расширить бронхи, кровеносные сосуды, зрачок.

Гладкая мышечная ткань называется неисчерченной, так как не обладает поперечной исчерченностью, характерной для поперечнополосатых скелетной и сердечной мышечных тканей.
Скелетная (поперечнополосатая) мышечная ткань
Скелетная мышечная ткань образует диафрагму (дыхательную мышцу), мускулатуру туловища, конечностей, головы, голосовых связок.
В отличие от гладкой мускулатуры, скелетная образована не отдельными одноядерными клетками, а длинными многоядерными волокнами, имеющими до 100 и более ядер — миосимпластами. Миосимпласт (греч. sim — вместе + plast — образованный) представляет совокупность слившихся клеток, имеет длину от нескольких миллиметров до нескольких сантиметров (соответствует длине мышцы).
Внутри миосимпласта находится саркоплазма, снаружи миосимпласт покрыт сарколеммой. Сократительные элементы — миофибриллы (лат. fibra — волоконце) — длинные тяжеобразные органеллы в миосимпласте (около 1400).

Характерная черта данной ткани — поперечная исчерченность, выражающаяся в равномерном чередовании светлых и темных полос на мышечном волокне. Это происходит потому, что границы саркомеров в соседних миофибриллах совпадают, вследствие чего все волокно приобретает поперечную исчерченность. Теперь самое время изучить микроскопическую основу мышцы — саркомер.
Саркомер (от греч. sarco — мясо (мышца) + mere — маленький)
Саркомер — элементарная сократительная единица поперечнополосатых мышц, структурная единица миофибриллы. В состав саркомера (и миофибриллы в целом) входят миофиламенты (лат. filamentum — нить) двух типов, которые обеспечивают сократимость мышечной ткани.
Саркомер состоит из актиновых (тонких) и миозиновых (толстых) филаментов, которые образованы главным образом белками актином и миозином. Сокращение происходит за счет взаимного перемещения миофиламентов: они тянутся навстречу друг другу, саркомер укорачивается (и мышца в целом).

Источником энергии для сокращения служат молекулы АТФ. К тому же невозможно представить сокращение мышц без участия ионов кальция: именно они связываются с тропонином, что приводит к изменению конформации тропомиозина (тропонин и тропомиозин — регуляторные белки между нитями актина), за счет чего становится возможно соединение актина и миозина. При сокращении мышц выделяется тепло (сократительный термогенез).
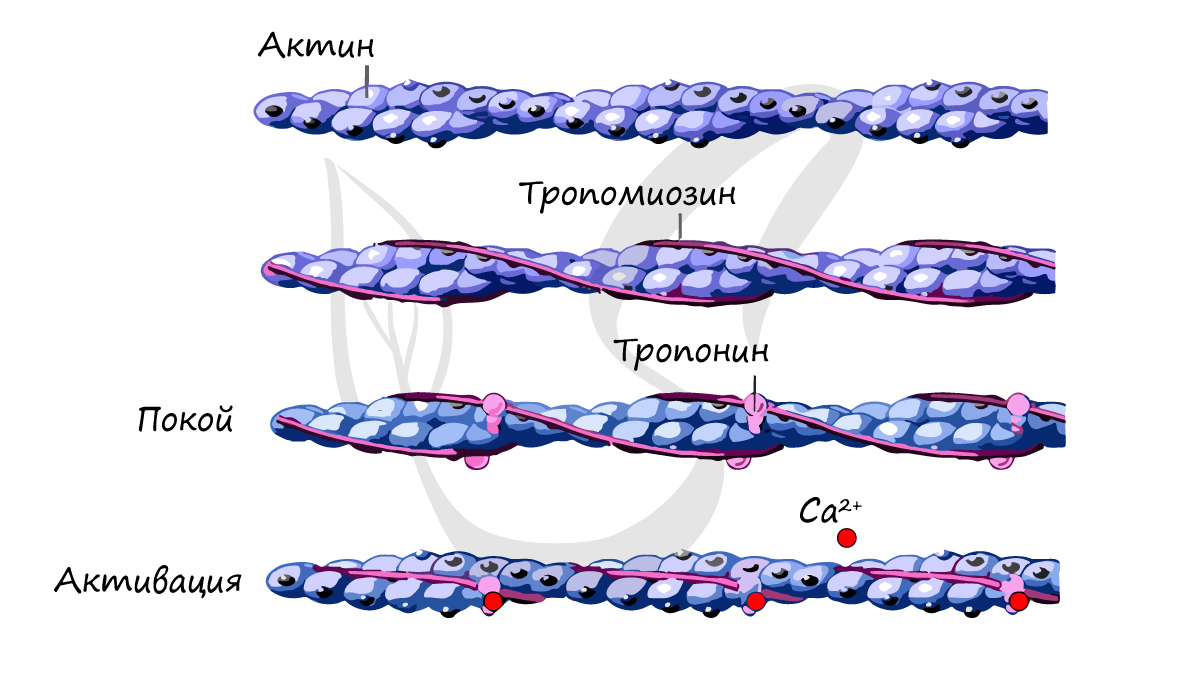
Замечу, что трупное окоченение (лат. rigor mortis) — посмертное затвердевание мышц — связано именно с ионами кальция, которые устремляются в область низкой концентрации (в саркоплазму миосимпласта), способствуя связыванию актина и миозина.
После смерти в мышце перестает синтезироваться АТФ, ее уровень быстро снижается. Как следствие этого перестает функционировать Ca-АТФаза — насос, выкачивающий ионы Ca из саркоплазмы в саркоплазматический ретикулум (мембранная органелла мышечных клеток (сходная с ЭПС), в которой запасаются ионы Ca).
В саркоплазме повышается концентрация ионов Ca — замыкаются мостики между актином и миозином, однако разомкнуться они уже не могут, в связи с чем наблюдается стойкая мышечная контрактура (лат. contractura — стягивание, сужение): конечности очень сложно разогнуть или согнуть.

Вернемся к скелетным мышцам. Имеется еще ряд важных моментов, о которых нужно знать.
Читайте также: Давление углекислого газа в тканях выше чем в крови или
В процесс возбуждения вовлекается изолированно один миосимпласт, соседние миосимпласты (волокна) не возбуждают друг друга, в отличие от гладких миоцитов, где возбуждение предается между соседними клетками через нексусы. Скелетные мышцы сокращаются быстро и быстро утомляются (у гладких мышц фазы сокращения и расслабления растянуты во времени, мало утомляются) .
Скелетные мышцы сокращаются произвольно: они подконтрольны нашему сознанию. К примеру, по желанию мы можем изменить скорость движения руки, темп бега, силу прыжка. Мышцы покрыты фасцией, крепятся к костям сухожилиями, и, сокращаясь, приводят в движение суставы.

Сердечная поперечнополосатая мышечная ткань
Сердечная мышечная ткань образует мышечную оболочку сердца — миокард (от др.-греч. μῦς «мышца» + καρδία — «сердце»). Миокард — средний слой сердца, составляющий основную часть его массы. При работе сердечная мышечная ткань не утомляется.

Сердечная мышечная ткань состоит из кардиомиоцитов — одиночных клеток, имеющих поперечную исчерченность. Соединяясь друг с другом, кардиомиоциты образуют функциональные волокна.
Этот тип мышечной ткани удивительным образом сочетает свойства двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство — автоматизм.
Автоматизм — способность сердечной мышечной ткани возбуждаться и сокращаться самопроизвольно, без влияний извне. Это легко можно подтвердить, наблюдая сокращения изолированного сердца лягушки в физиологическом растворе: сокращения сердца в нем будут продолжаться несколько десятков минут после отделения сердца от организма.

Места контактов соседних кардиомиоцитов — вставочные диски (в их составе находятся нексусы), благодаря которым возбуждение одной клетки передается на соседние, таким образом волнообразно охватываются возбуждением и сокращаются новые участки миокарда.
Большое число контактов между кардиомиоцитами обеспечивает высокую эффективность и надежность проведения возбуждения по миокарду. Сокращается эта ткань непроизвольно, не утомляется.
На рисунке или микропрепарате узнать данную ткань можно по центральному положению ядер в клетках, поперечной исчерченности, наличию вставочных дисков и анастомозов (греч. anastomosis — отверстие) — мест соединений боковых поверхностей функциональных волокон (кардиомиоцитов).

В норме возбуждение проводится по проводящей системе сердца от предсердий к желудочкам (однонаправленно). Участок сердечной мышцы, в котором генерируются импульсы, определяющие частоту сердечных сокращений — водитель сердечного ритма.
Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных (англ. pacemaker — задающий ритм) клеток, которые также называют водителями ритма. Они спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.
Ответ мышц на физическую нагрузку
Физические нагрузки приводят к гипертрофии мышц (от др.-греч. ὑπερ- чрез, слишком + τροφή — еда, пища) — в них увеличивается количество мышечных волокон, объем мышечной массы нарастает.

В условиях гиподинамии (от греч. ὑπό — под и δύνᾰμις — сила), то есть пониженной активности, мышцы уменьшаются вплоть до полной атрофии (греч. а – «не» + trophe – питание). В худшем случае волокна мышечной ткани перерождаются в соединительную ткань, после чего пациент становится обездвиженным.

Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление. Гипертрофия сердца — состояние, требующее вмешательства врача и наблюдения за пациентом.
В большинстве случае гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы).

Происхождение мышц
Мышцы развиваются из среднего зародышевого листка — мезодермы.

© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Мышечные ткани
Св-вом сократимости обладают практически все виды кл-к, благодаря наличию в их цитоплазме сократительного аппарата, представленного микрофиламентами, состоящими из сократительных белков – актина, миозина и др. Наиболее выражены сократительные процессы в клетках, основной ф-цией которых является сокращение. Такие клетки или их производные образуют мышечные ткани, которые обеспечивают сократительные процессы в полых внутренних органах и сосудах, перемещение частей тела относительно друг друга, поддержание позы и перемещение организма в пространстве. Помимо движения при сокращении выделяется большое количество тепла, а, следовательно, мышечные ткани участвуют в терморегуляции. Мышечные ткани неодинаковы по строению, источникам происхождения и иннервации, по функциональным особенностям.
Читайте также: Зачем кусочки ткани для одежде
КЛАССИФИКАЦИЯ МЫШЕЧНЫХ ТКАНЕЙ
Обе разновидности развиваются из мезодермы, но из разных её частей: скелетная – из миотомов сомитов, сердечная – из висцерального листка спланхнотома.
— мезенхимного происхождения: мышечная ткань сосудов и внутренних органов
— эктодермального происхождения: миоэпителиоциты потовых, молочных, слюнных и слёзных желез
— мионейрального происхождения: мышцы суживающие и расширяющие зрачок.
Каждая разновидность мышечной ткани имеет свою структурно- функциональную единицу. Структурно-функциональной единицей гладкой мышечной ткани внутренних органов и радужной оболочки является гладкий миоцит, у мышечной ткани эктодермального происхождения – корзинчатый миоэпителиоцит, сердечной мышечной ткани – кардиомиоцит, у скелетной мышечной ткани – мышечное волокно.
ПОПЕРЕЧНО-ПОЛОСАТАЯ СКЕЛЕТНАЯ МЫШЕЧНАЯ ТКАНЬ
П.с. цилиндрическое образование с заострёнными концами, диаметром около 100 мкм. Состоит из 2 частей:
1) симпластическая часть (симпласт)
Симпласт снаружи покрыт сарколеммой, которая состоит из 2 листков:
— внутренний – типичная плазмолемма
— наружный – тонкая соединительнотканная базальная пластинка.
Между базальной пластинкой и плазмолеммой находятся миосателлиты – камбиальные элементы. За счёт них осуществляется регенерация мышечного вол-на.
В миосимпласте содержится несколько тысяч ядер, располагающихся на периферии под плазмолеммой. Органеллы общего назначения развиты слабо, центриоли отсутствуют. В саркоплазме содержатся включения гликогена и миоглобина, аналога гемоглобина эритроцитов. Отличительной особенностью симпласта является наличие органелл специального назначения, предназначенных для выполнения сократительной ф-ции:
2) саркоплазматическая сеть
Основную часть миосимпласта занимают миофибриллы, которые локализуются в его центральной части. Каждая миофибрилла простирается на протяжении всего миосимпласта и своими свободными концами прикрепляется к его плазмолемме у конических концов. Между миофибриллами располагаются митохондрии – саркосомы. Миофибриллы неоднородны по строению и состоят из тёмных (анизотропных) А-дисков и светлых (изотропных) И-дисков. Тёмные и светлые диски всех миофибрилл располагаются на одном уровне и обуславливают поперечную исчерченность всего мышечного вол-на. Темные и светлые диски состоят из более тонких волоконец – миофиламентов. Тёмные диски образованы толстыми миофиламентами (10-12 нм), состоящими из белка миозина. Светлые диски образованы тонкими миофиламентами (5-7 нм), состоящими из белка актина. Актиновые миофиламенты свободными концами частично входят в А-диск между толстыми миофиламентами. При этом вокруг одного миозинового филамента располагаются 6 актиновых. Посередине И-диска проходит полоска, которая называется зет-линия (телофрагма), посередине А – диска – тёмная М=линия – мезофрагма. Участок миофибриллы, лежащий между 2 соседними зет-линиями, называется саркомером. Саркомер – это структурно-функциональная единица миофибриллы. В состав саркомера входят: зет-линия, одна вторая диска И. диск А, одна вторая диска И, вторая зет-линия. Каждая миофибрилла п.с. совокупность саркомеров.
Для реализации сокращения необходимы 3 условия:
наличие энергии в виде АТФ
АТФ образуется в саркосомах . Выполнение 2 последних условий возможно благодаря наличию саркоплазматической сети и Т-канальцев.
Саркоплазматическая сеть – это видоизменённая гл. ЭПС. Подразделяется на фрагменты, окружающие отдельные саркомеры. Каждый фрагмент состоит из 2 терминальных цистерн, соединённых полыми анастомозирующими Л-канальцами. Терминальные цистерны охватывают саркомер в области И-дисков, канальцы – в области А-дисков.
В саркоплазматической сети содержатся ионы кальция, которые при распространении нервного импульса выходят из цистерн и канальцев и инициируют взаимодействие актиновых и миозиновых миофиламентов.
Волна деполяризации передаётся от нервного окончания на саркоплазматическую сеть через Т-канальца. Они п.с. впячивания плазмолеммы в саркоплазму. Проникая вглубь. Т-канальца разветвляются и охватывают каждую миофибриллу обычно на уровне зет-полоски. По сторонам от каждого Т-канальца располагаются 2 терминальные цистерны саркоплазматической сети. Эта структура носит название триады.
МЕХАНИЗМ МЫШЕЧНОГО СОКРАЩЕНИЯ
Общепринятой теорией является модель сокращения Х. Хаксли или теория скольжения нитей.
Суть: нервный импульс передаётся на плазмолемму мышечного вол-на. Затем возбуждение идет на Т-трубочки и распространяется на лежащие рядом терминальные цистерны. Из цистерн СПР выходят ионы кальция. Основной точкой их приложения являются актиновые филаменты. На них кальций открывает активные центры для связывания головок миозина.
Процесс сокращения осуществляется благодаря взаимодействию актиновых и миозиновых филаментов и образованию между ними акто-миозиновых мостиков.
За счёт укорочения этих мостиков происходит втягивание актиновых филаментов между миозиновыми.
Затем эти связи распадаются и головки миозина образуют новые контакты с другими точками на актиновом филаменте, но расположенными дистальнее предыдущих. Так происходит укорочение саркомера.
При частичном сокращении миофибриллы в А-диске образуется светлая зона или Н-полоска, ограниченная свободными концами актиновых филаментов. Ширина Н-полоски зависит от степени сокращения миофибриллы. Формула саркомера может быть представлена в следующем виде:
При полном сокращении саркомера актиновые филаменты достигают М-полоски саркомера. При этом исчезает полоска и И-диски и формула саркомера выглядит следующим образом:
Читайте также: Травма мягких тканей что это
Структурные элементы саркомера в расслабленном состоянии можно выразить формулой:
Различают 2 основных типа мышечных волокон:
вол-на 1 типа – красные мышечные вол-на – хар-ся высоким содержанием в саркоплазме миоглобина, что и придает им красный цвет, большим числом саркосом, высокой активностью окислительно-восстановительных ф-тов. Эти вол-на способны к медленному, но длительному тоническому сокращению. Отличаются малой утомляемостью.
вол-на 2 типа – хар-ся незначительным содержанием миоглобина, но высоким содержанием гликогена и высокой активностью гликолитических ф-тов. Способны к быстрому, сильному, но непродолжительному сокращению.
Между крайними типами находятся промежуточные мышечные вол-на, занимающие и в структурном и в функциональном отношении промежуточное положение между первыми двумя.
Мышца как орган — это анатомическое образование, состоит из мышечных вол-н, волокнистой соединительной ткани, сосудов и нервов.
Волокнистая соединительная ткань образует в мышце прослойки: эндомизий, перимизий и эпимизий. Эндомизий окружает каждое мышечное вол-но, содержит кровеносные и лимфатические сосуды, посредством которых обеспечивается трофика мышечного вол-на.. Перимизий окружает несколько мышечных вол=н, собранных в пучки. Эпимизий или фасция окружает всю мышцу, способствует функционированию мышцы, как органа.
СЕРДЕЧНАЯ ПОПЕРЕЧНО-ПОЛОСАТАЯ МЫШЕЧНАЯ ТКАНЬ.
Стуруктурно-функциональной единицей является кардиомиоцит. Различают 3 типа кардиомиоцитов:
Атипичные (проводящие) кардиомиоциты.
Рабочие (сократительные) кардиомиоциты имеют прямоугольную форму. Снаружи покрыты базальной пластинкой. Миосателлиты отсутствуют, поэтому регенерация возможна только заместительным способом – образованием рубца. Ядро обычно 1, локализовано в центре. Периферическую часть саркоплазмы занимают миофибриллы, а между ними в большом кол-ве локализуются митохондрии. Миофибриллы кардиомиоцитов анастомозируют друг с другом, образуя сеть, поэтому поперечная исчерченность выражена не отчётливо. СПР представлена расширенными канальцами. Терминальные цистерны и триады отсутствуют. Т-трубочки короткие и широкие.
Место контакта 2 кардиомиоцитов наз-ся вставочным диском. Вставочный диск – это комплекс межклеточных контактов, в котором различают 3 зоны:
Посредством вставочных дисков обеспечивается механическая и функциональная связь кардиомиоцитов. Наличие нексусов обеспечивает одновременное и содружественное сокращение кардиомиоцитов вначале в предсердиях, затем в желудочках.
Соединяясь друг с другом, кардиомиоциты образуют функциональные мышечные вол-на. Между ними имеются анастомозы, благодаря которым обр-ся сеть – функциональный синцитий.
Секреторные кардиомиоциты. Локализуются в основном в правом предсердии. Вырабатывают натрийуретический и антидиуретический факторы. Эти вещ-ва влияют на уровень давления крови в сердце и сосудах , и препятствуют образованию тромбов в предсердиях.
Проводящие (атипичные) кардиомиоциты – образуют проводящую систему сердца:
— пучок Гиса – ствол, правая и левая ножки
— волокна Пуркинье – концевые разветвления ножек пучка гиса.
Атипичные кардиомиоциты обеспечивают генерирование, проведение и передачу на сократительные кардиомиоциты нервных импульсов.
Особенности морфологии атипичных кардиомиоцитов:
— в цитоплазме мало миофибрилл, расположены они неупорядоченно, поэтому атипичные кардиомиоциты не имеют поперечной исчерченности.
— вставочные диски отсутствуют
— в саркоплазме выявляется большое кол-во гликогена.
Атипичные кардиомиоциты различаются между собой по структуре и ф-циям и подразделяются на 3 основные разновидности:
Р-кл-ки – пейсмекеры, водители ритма. Составляют основу синусно- предсердного узла, с определённой частотой способны генерировать нервные импульсы и передавать их на кл-ки 2 типа
2 тип – переходные клетки – содержатся в предсердно-желудочковом узле.
3 тип – кл-ки пучка Гиса и вол-н пуркинье. – передают импульсы на сократительные кардиомиоциты.
Структурно-функциональная единица — гладкий миоцит. Он п.с. веретенообразную клетку, покрытую базальной пластинкой. В центре располагается палочковидное ядро, по полюсам которого локализуются органеллы общего назначения. В цитоплазме содержатся актиновые и миозиновые филаменты, которые располагаются параллельно друг другу и не образуют А и И –дисков. Этим объясняется отсутствие поперечной исчерчености миоцитов. Плазмолемма образует небольшие углубления – кавеолы, которые являются аналогами Т-трубочек. В цитоплазме локализуются многочисленные везикулы, которые являются элементами СПР.
В цитоплазме и на периферии миоцитов под их плазмолеммой локализуются плотные тельца, к которым прикрепляются актиновые и миозиновые филаменты.
Механизм сокращения осуществляется за счёт взаимодействия и скольжения актиновых филаментов вдоль миозиновых. Для такого взаимодействия также необходимы энергия в виде АТФ, ионы кальция и наличие биопотенциала. Головки молекул миозина взаимодействуют с активными центрами актиновых филамент и совершают тянущие гребковые движения. В результате этих гребковых движений сближаются плотные тельца, и гладкий миоцит сокращается.
Гладкие миоциты функционируют не изолированно, а формируют миоцитарные комплексы, которые состоят из 10-12 миоцитов. В составе комплекса миоциты взаимодействуют друг с другом при помощи десмосом и нексусов. В области нексусов базальные мембраны миоцитов прерываются, происходит передача возбуждения от одного миоцита к соседним, в результате чего сокращением охватывается весь комплекс.
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом