
Осмотический потенциал растительной ткани составляет
Чтобы понять, как осуществляется поддержание водного режима растений, необходимо разобраться в таких процессах, как осмос и диффузия, которые рассмотрены в нашей статье. Там подчеркнуто, что осмос можно считать типом диффузии, при которой диффундируют только молекулы воды, а молекулы всех остальных веществ не проходят через полупроницаемую мембрану. Молекулы воды перемещаются через полупроницаемую мембрану из области с высокой их концентрацией (т. е. из относительно разбавленного раствора) туда, где она ниже (т. е. в более концентрированный раствор).
В 1988 г. Институт биологии рекомендовал пользоваться термином водный потенциал при описании движения воды через мембраны. Водный потенциал растительных клеток определяется двумя главными факторами — концентрацией растворенных веществ и давлением, возникающим при набухании клетки в результате поступления в нее воды. Соответственно говорят о концентрационном, или осмотическом, и гидростатическом потенциалах. Эти термины объясняются ниже.

Водный потенциал
Водный потенциал — один из фундаментальных показателей термодинамического состояния воды. Ее молекулы обладают кинетической энергией, поскольку и в жидкости, и в газе (водяном паре) быстро и беспорядочно движутся. Чем выше концентрация этих молекул в системе, тем больше их общая кинетическая энергия и выше водный потенциал. Таким образом, у чистой воды он максимален. Если две системы, содержащие воду (например, почва и атмосфера или клетка и окружающий ее раствор), вступают в контакт, то беспорядочное движение ее молекул приведет к их результирующему движению туда, где их меньше, — из области с более высоким водным потенциалом (с более высокой энергией) туда, где он ниже (ниже энергия), пока их концентрации в обеих системах не уравновесятся.
В биологии водный потенциал обычно измеряют единицами давления (например, Паскалями)1. У чистой воды он максимален, но условно принят за нулевой.
Запомните следующие главные моменты:
1) водный потенциал чистой воды максимален и, по определению, равен нулю (в «стандартных» условиях, т. е. при нормальном атмосферном давлении и 25 °С);
2) вода всегда движется по градиенту у: оттуда, где он выше, туда, где он ниже;
3) у всех растворов водный потенциал ниже, чем у чистой воды, т. е. в стандартных условиях выражается отрицательной величиной;
4) осмос можно определить как движение молекул воды через полупроницаемую мембрану по градиенту водного потенциала.
Преимущество использования понятия «водный потенциал»
Водный потенциал отражает тенденцию молекул воды двигаться в определенном направлении. Чем он выше (чем менее отрицателен), тем больше у молекул шансов покинуть данную систему. Если две системы (не обязательно разделенные мембраной) контактируют друг с другом, то вода будет переходить из системы с более высоким водным потенциалом в систему с более низким водным потенциалом.
Таким образом, понятие «водный потенциал» позволяет определить направление движения воды между любыми двумя системами — не только из одной клетки растения в другую, но, например, из почвы в корень, из листа в атмосферу и т. п. Можно сказать, что вода движется через растение по градиенту водного потенциала из почвы в воздух. Чем круче этот градиент, тем быстрее поток воды.
5.Осмотический потенциал растительной ткани ,, методы определения и возможност использования в сельскохозяйственной практике.
Односторонняя диффузия молекул воды или другого растворителя через полупроницаемую мембрану называется осмос. Причиной осмоса является разность концентрации растворов по обе стороны полупроницаемой мембраны. В 1748 А. Ноллет наблюдал, как растворитель проходит через мембрану из разбавленного раствора в более концентрированный.
Система, в которой можно наблюдать осмос, называется осмотической. Она состоит из растворов разных концентраций или из раствора и растворителя, разделенных полупроницаемой мембраной. Пространство, окруженное такой мембраной и заполненное каким-нибудь раствором получило, название осмотической ячейки.
Изучение осмоса в растительной клетке началось давно в 1826г. Французский ботаник Г. Дютроше смастерил: к кончику стеклянной трубки он привязал мешочек из пергамента, заполненный раствором соли или сахара, и опускал его в стакан с водой. При этом вода поступала в мешочек и раствор немного поднимался по трубке.
В 1877 г. Немецкий ботаник В. Пфеффер создал более совершенную модель растительной клетки, названную осмометром Пфеффера.
В цитоплазме находится очень много разных органелл, окруженных мембранами, то всю ее в данном случае тоже можно считать полупроницаемой. Любая органелла цитоплазмы, окруженная мембраной, представляет собой осмотическую ячейку. Осмотическое передвижение воды происходит также между отдельной органеллой и цитозолем. Идеальная полупроницаемая мембрана пропускает молекулы воды и не пропускает молекул растворенного вещества. Сопротивление движению воды зависит от липидного бислоя и от строения и расположения белковых глобул.
Читайте также: Крой постельного белья из ткани шириной 150 см
Дополнительное давление, которое необходимо приложить к раствору, чтобы помешать одностороннему току растворителя (воды) в раствор через полупроницаемую мембрану, назвали осмотическим давлением. К веществам осмотически активными относятся органические кислоты, аминокислоты, сахара, соли. Раствор, имеющий большее осмотическое давление, называется гипертоническим, меньшее – гипотоническим.
Поступая в клетку, вода тем самым увеличивает объем вакуоли, которая давит на цитоплазму и заставляет протопласт прижиматься к клеточной стенке. Клеточная стенка растягивается, отчего клетка переходит в напряженное состояние – тургор. Давление протопласта на клеточную стенку получило название тургорное. Так как клеточная стенка имеет небольшую эластичность, она начинает давить на протопласт в противоположном направлении. Это давление клеточной стенки на протопласт называется тургорным натяжением.
Осмотическое поступление воды приводит к возникновению тургорного давления. Разница между осмотическим давлением клеточного сока и противодавлением клеточной стенки определяет поступление вод в клетку каждый данный момент.
Химический потенциал воды называется водным потенциалом. Чем ниже энергия молекул воды, тем ниже водный потенциал. Химический потенциал любого раствора – отрицательная величина и с увеличением концентрации растворенных веществ становится все более отрицательным.
Водный потенциал клетки – разность между свободной энергией воды внутри и вне клетки при той же температуре и атмосферном давлении. Величина водного потенциала клетки определяется степенью ее насыщенности водой: чем больше клетка насыщена водой, тем менее отрицателен ее водный потенциал.
Компоненту водного потенциала клетки, которая определяется присутствием растворенного вещества, обозначают особым термином – «осмотический потенциал». Осмотический потенциал раствора связан прямой зависимостью с концентрацией растворенного вещества. С увеличением этой концентрации осмотический потенциал становится все более отрицательной величиной.
Положительный потенциал тургорного давления полностью уравновешивает отрицательный осмотический потенциал, и клетка перестает поглощать воду; в таком состоянии ее водный потенциал равен нулю. Это состояние называется состоянием насыщения. Поскольку осмотический потенциал равен разности между химическим потенциалом раствора и химическим потенциалом чисто воды, который равен 0, то он всегда отрицательный.
Для клеток деревьев в состав этого уравнения включают еще одно слагаемое – гравитационный потенциал отражающий влияние на активность воды силы тяжести, заметно сказывающийся только при подъеме воды на большую высоту.
Величина осмотического потенциала изменяется и в пределах растения: у корней – 0,5 — — 1,0, у верхних листьев – до – 4,0 МПа. Это обуславливает существование градиента водного потенциала клеток от корней к листьям. Величина осмотического потенциала зависит также от температуры, интенсивности света.
Энергия для осмоса нужна: во-первых, нужно иметь растворы разной концентрации по обе стороны мембраны; энергия тратится на активный транспорт растворенных веществ в вакуоль и создание градиента концентрации. Во-вторых, осмотически активные вещества, накапливающиеся в вакуоли, является продуктами метаболизма, следовательно, для их образования тоже затрачивается энергия. В- третьих, энергия необходима для сохранения избирательной проницаемости мембран.
Осмотические процессы лежат в основе многих процессов, например поступления воды, движения органов растения, движения устьиц.
При достаточном водоснабжении вода в коре корня транспортируется главным образом по апопласту, т.к. здесь она встречает менее сопративление, и лишь частично по симпласту. Если клетки корня испытывают недостаток воды, то она будет предвигаться в основном по симпласту.
Живые клетки оказывают большее сопротивление движению воды, поэтому по ним передвигается лишь 1-10% ее общего количества.
В листовой пластинке транспорт воды идет в основном по апопласту. Транспорт воды в листовой пластинке изучен хуже, чем в корне.
Б.5)Осмотический потенциал растительной ткани, методы определения и возможности использования в сельскохозяйственной практике.
Растительная клетка представляет собой осмотическую систему. Пектоцеллюлозная оболочка хорошо проницаема как для воды, так и для растворенных веществ. Однако плазмалемма и тонопласт обладают избирательной проницаемостью, легко пропускают воду и менее проницаемы, а в некоторых случаях непроницаемы для растворенных веществ. В этом можно убедиться, рассмотрев явления плазмолиза и тургора. Если поместить клетку в раствор более высокой концентрации, чем в клетке, то под микроскопом видно, что цитоплазма отстает от клеточной оболочки. Это особенно хорошо проявляется на клетке с окрашенным клеточным соком. Клеточный сок остается внутри вакуоли, а между цитоплазмой и оболочкой образуется пространство, заполненное внешним раствором. Явление отставания цитоплазмы от клеточной оболочки получило название плазмолиза. Плазмолиз происходит в результате того, что под влиянием более концентрированного внешнего раствора вода выходит из клетки (от своего большего химического потенциала к меньшему), тогда как растворенные вещества остаются в клетке. При помещении клеток в чистую воду или в слабо концентрированный раствор вода поступает в клетку. Количество воды в клетке увеличивается, объем вакуоли возрастает, клеточный сок давит на цитоплазму и прижимает ее к клеточной оболочке. Под влиянием внутреннего давления клеточная оболочка растягивается, в результате клетка переходит в напряженное состояние — тургор.
Читайте также: Микро поплин что за ткань
Наблюдения за явлениями плазмолиза и тургора позволяют изучить многие свойства клетки. Явление плазмолиза показывает, что клетка жива и цитоплазма сохранила полупроницаемость. В мертвых клетках мембрана не обладает полупроницаемостью, не контролирует потоки веществ, и осмотический выход воды не происходит. По скорости и форме плазмолиза можно судить о вязкости цитоплазмы. Наконец, явление плазмолиза позволяет определить величину осмотического потенциала в клетке (плазмолитический метод). Этот метод основан на подборе изоосмотического, или изотонического, раствора, т. е. имеющего осмотический потенциал (Ψосм. р-ра) , равный осмотическому потенциалу клеточного сока (Ψосм. кл.). Раствор, при котором в клетке начался плазмолиз, имеет осмотический потенциал, примерно равный осмотическому потенциалу клетки. Зная концентрацию этого наружного раствора в молях, можно вычислить его осмотический потенциал, а следовательно, осмотический потенциал клетки (Ψосм. р-ра = Ψосм. кл.).
Определение величины осмотического потенциала имеет большое значение, в частности для экологических исследований. Величина осмотического потенциала позволяет судить о максимальной способности растения поглощать воду из почвы и удерживать ее, несмотря на иссушающее действие атмосферы. Осмотический потенциал колеблется в широких пределах, от —5 до —200 бар. Осмотический потенциал около —1 бара наблюдается у водных растений. Осмотический потенциал, равный —200 бар, обнаружен у выжатого сока талофтаAtriplexconfertifolia. В 1 л сока этого растения содержится 67,33 г хлоридов. У большинства растений средней полосы осмотический потенциал колеблется от —5 до —30 бар. Вместе с тем необходимо отметить, что факторы, действующие на изменение осмотического потенциала, чрезвычайно разнообразны. Даже соседние, рядом расположенные клетки могут отличаться по величине осмотического потенциала. Обычно отрицательная величина осмотического потенциала больше у мелких клеток по сравнению с крупными. Установлены определенные градиенты осмотического потенциала в пределах одной ткани. Так, в тканях стебля отрицательный осмотический потенциал возрастает от периферии к центру и от основания к верхушке. В корне отрицательный осмотический потенциал, наоборот, постепенно снижается от основания к верхушке. В проводящих элементах стебля и корня, как правило, отрицательная величина осмотического потенциала очень низка (от —1 до —1,5 бара). В листьях осмотический потенциал колеблется от -10 до -18 бар. Осмотический потенциал различен у разных жизненных форм. У древесных Пород он более отрицателен, чем у кустарников, а у кустарников более отрицателен, чем у травянистых растении. Разные экологические группы различаются по величине осмотического потенциала. У растений пустынь осмотический потенциал более отрицателен, чем у степных растений; у степных более отрицателен, чем у луговых. Еще меньше осмотическая концентрация у растений болотных и водных местообитаний (соответственно наименее отрицательный осмотический потенциал). У светолюбивых растений осмотический потенциал более отрицательный, чем у теневыносливых. На величину осмотического потенциала влияет концентрация растворенных веществ в клеточном соке — это осмотически активные вещества (органические кислоты, соли, аминокислоты, сахара). Растение в определенной степени регулирует величину осмотического потенциала. Ферментативное превращение сложных нерастворимых веществ в растворимые (крахмала в сахара, белков в аминокислоты) приводит к возрастанию концентрации клеточного сока и повышению отрицательной величины осмотического потенциала. Увеличенное накопление растворимых солей также делает более отрицательным осмотический потенциал. Несмотря на то, что осмотический потенциал меняется в зависимости от внешних условий, все же для каждого вида эти изменения происходят в своих определенных пределах. Величину осмотического потенциала многие физиологи считают одной из характеристик данного вида растений.
Осмотический потенциал растительной ткани составляет
При растворении в воде другого вещества, концентрация ее молекул, а, следовательно, и водный потенциал, снижаются. Таким образом, у всех растворов водный потенциал ниже, чем у чистой воды. Количественно это понижение выражают величиной, называемой осмотическим потенциалом (у0). Иными словами, осмотический потенциал — это мера снижения водного потенциала системы в результате присутствия в ней «неводных» молекул. Осмотический потенциал водного раствора всегда отрицателен. Чем больше в растворе молекул растворенного вещества, тем этот потенциал ниже.
Читайте также: Гост показатели тканей для спецодежды
Гидростатический потенциал
Если к чистой воде или раствору приложить давление, то водный потенциал возрастает, поскольку у жидкости возникает тенденция переместиться в другое место. Такая ситуация возможна в живой клетке. Например, когда за счет осмоса в нее поступает вода, клетка набухает, и внутри нее повышается давление, называемое тургорным. Сходным образом водный потенциал плазмы крови повышается до положительной отметки кровяным давлением в почечных клубочках. Гидростатический потенциал обычно положителен, но в некоторых случаях, например когда столб воды, «висящий» в ксилеме, растягивается, он может стать отрицательным (возникает отрицательное давление).
На водный потенциал влияет как концентрация растворенного в воде вешества, так и давление, поэтому можно выразить его в виде уравнения:
Водный потенциал = Осмотический потенциал + Гидростатический потенциал
Осмотический потенциал всегда отрицателен, а гидростатический обычно положителен.
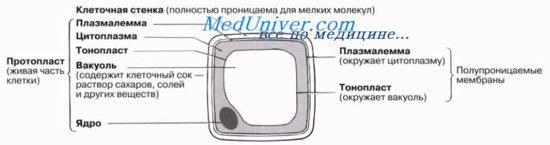
Осмос и растительные клетки
На рисунке показаны полупроницаемые мембраны, имеюшие важное значение для водного режима растительной клетки. Клеточная стенка обычно полностью проницаема для любых растворенных молекул, поэтому ее нельзя считать осмотическим барьером. В клетке часто имеется крупная центральная вакуоль, содержимое которой, клеточный сок, влияет на общий осмотический потенииап системы. В целом водный режим клетки зависит от двух мембран — плазмалеммы, окружающей снаружи цитоплазму, и тонопласта, ограничивающего вакуоль.
Если растительная клетка контактирует с раствором, водный потенци&т которого ниже, чем у ее содержимого (например, с концентрированным сахарным сиропом), то вода будет выходить из нее за счет осмоса через плазмалемму. Сначала воды станет меньше в цитоплазме, а затем и в вакуоле, откуда она выйдет сквозь тонопласт. Протопласт, т. е. живое содержимое растительной клетки, окруженное клеточной стенкой, спадется и отойдет от этой стенки, как бы сжавшись внутри нее. Этот процесс называется плазмолизом, а клетка в таком состоянии — плазмолизированной. Момент, когда протопласт еще прилегает к клеточной стенке, но уже перестал оказывать на нее давление, называется начальным плазмолизом. В этот момент клетка теряет тургор, т. е. становится вялой. Вода будет покидать протопласт до тех пор, пока его содержимое по водному потенциалу не сравняется с окружающим раствором. Тогда установится равновесное состояние, и спадение протопласта прекратится.
Обычно плазмолиз обратим и не причиняет существенного вреда клетке. Если затем перенести клетку в чистую воду или в раствор с более высоким водным потенциалом, чем у цитоплазмы, то вода будет поступать в нее за счет осмоса. По мере увеличения объема протопласта он станет давить на клеточную стенку и растянет ее. Эта стенка прочная и относительно жесткая, поэтому давление на нее быстро возрастет — повысится гидростатический (\\/г) потенциал клетки. Внешне поступление в нее воды путем осмоса приведет к набуханию, напряженному состоянию, которое называется тургором. Говорят, что клетка становится тургесцентной, а давление друг на друга ее содержимого и стенки называют тургорным. Полная тургесцентность, т. е. максимальное значение \\/г, достигаются при погружении клетки в чистую воду.
Когда водный потенциал окружающего клетку раствора уравновесится ее возросшим гидростатическим потенциалом, внутрь будет проникать столько же водных молекул, сколько выходить наружу за то же время. Несмотря на продолжающееся их движение через мембрану, изменения системы в целом прекратятся. Такое равновесие со средой называется динамическим. Осмотический потенциал клеточного содержимого останется скорее всего ниже, чем снаружи, поскольку для роста тургорного давления нужно не так много воды — меньше, чем для существенного разбавления внутреннего раствора. Однако эта разница компенсируется более высоким, чем снаружи, гидростатическим потенциалом клетки. Суммарные же водные потенциалы с обеих сторон клеточной стенки сравняются.
Тургорное давление может вырасти только в замкнутом объеме, ограниченном клеточной стенкой. У животных клеток такой стенки нет, а их наружная мембрана слишком тонка, чтобы сдержать набухание цитоплазмы при поступлении внутрь воды из раствора с более высоким водным потенциалом. В такой среде они просто сильно набухнут и лопнут, если не будут защищены особым механизмом осморегуляции.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом