
Основная функция гладкой эпс в волокнах поперечно полосатых мышечных тканей
Это ткань энтомезенхимного происхождения, которая делится на два вида: висцеральную и сосудистую. В эмбриональном гистогенезе даже электронно-микроскопически трудно отличить мезенхимные предшественники фибробластов от гладких миоцитов. В малодифференцированных гладких миоцитах развиты гранулярная эндоплазматическая сеть, комплекс Гольджи. Тонкие филаменты ориентированы вдоль длинной оси клетки. По мере развития размеры клетки и число филаментов в цитоплазме возрастают. Постепенно объем цитоплазмы, занятый сократительными филаментами, увеличивается, расположение их становится все более упорядоченным. Пролиферативная активность гладких миоцитов в миогенезе постепенно снижается. Это происходит в результате увеличения продолжительности клеточного цикла, выхода клеток из цикла репродукции и перехода в дифференцированное состояние.
Однако и в дефинитивном состоянии в гладкой мышечной ткани клеточная регенерация в виде размножения миоцитов полностью не прекращается. Существуют данные о том, что пролиферация и дифференцировка в большей степени свойственна субпопуляции малых (по размерам) гладких миоцитов.
Строение гладкой мышечной ткани. Структура дефинитивных гладких миоцитов (лейомиоцитов), входящих в состав внутренних органов и стенки сосудов, имеет много общего, но в то же время характеризуется гетероморфией. Так, в стенках вен и артерий обнаруживаются овоидные, веретеновидные, отростчатые миоциты длиной 10-40 мкм, доходящие иногда до 140 мкм.
Гладкая мышечная ткань
Наибольшей длины гладкие миоциты достигают в стенке матки — до 500 мкм. Диаметр миоцитов колеблется от 2 до 20 мкм. В зависимости от характера внутриклеточных биосинтетических процессов различают контрактилъные и секреторные миоциты. Первые специализированы на функции сокращения, но вместе с тем сохраняют секреторную активность. Плазмолемма расслабленной клетки имеет ровную поверхность, а при сокращении становится складчатой. В центре клетки имеется палочковидное ядро, которое при сокращении клетки спиралевидно изгибается. Практически все ядра миоцитов содержат диплоидное количество ДНК. Гладкая эндоплазматическая сеть занимает примерно 2-7% объема цитоплазмы, а гранулярная сеть в контрактильных миоцитах выражена плохо. Митохондрии мелкие, сферические или овоидные, расположены у полюсов ядра. Характерной чертой гладких миоцитов является наличие множества впячиваний (кавеол) плазмолеммы, содержащих ионы кальция.
Секреторные миоциты (синтетические) по своей ультраструктуре напоминают фибробласты, однако содержат в цитоплазме пучки тонких миофиламентов, расположенные на периферии клетки. В цитоплазме хорошо развиты комплекс Гольджи, гранулярная эндоплазматическая сеть, много митохондрий, гранул гликогена, свободных рибосом и полисом. По степени зрелости такие клетки относят к малодифференцированным.
Сократительный аппарат миоцитов представлен тонкими актиновыми филамен-тами (гладкомышечным альфа-актином), связанными с тропомиозином. Толстые нити состоят из миозина, мономеры которого располагаются вблизи филаментов актина. Соотношение актиновых и миозиновых филаментов в гладком миоците составляет 12 к 1. Важным компонентом контрактильного аппарата миоцитов являются электронно-плотные структуры — тельца прикрепления, расположенные свободно в цитоплазме (плотные тельца) или тесно связанные с плазмолеммой. Основными белковыми компонентами плотных телец являются альфа-актинин, актин (немышечный) и кальпонин, что позволяет расссматривать их как функциональный эквивалент Z-линий миофибрилл скелетной мышцы. Актиновые филаменты фиксируются на плотных тельцах. Промежуточные филаменты, включающие десмин и виментин, обеспечивают связи между плотными тельцами и плазмолеммой, образуя прикрепительные пластины.
Сократительные белки формируют решетчатую структуру, закрепленную по окружности плазмолеммы, поэтому сокращение выражается в укорочении клетки, которая приобретает складчатую форму, тогда как в состоянии покоя клетка вытянута. При возникновении нервного импульса, распространяющегося по плазмолемме миоцита, происходит повышение уровня внутриклеточного Са2+, который поступает в цитоплазму из кавеол, отшнуровывающихся в цитоплазму в виде пузырьков. Высвобождение ионов кальция приводит к каскаду реакций, в результате которого происходит полимеризация миозина и образование перекрестных связей миозина вдоль актиновых филаментов по мере развития мышечного сокращения. Расслабление мышцы возникает при восстановлении концентрации исходного уровня Са2+ внутри клетки путем его перемещения внутрь саркоплазматической сети. При этом образовавшиеся в присутствии ионов кальция связи между актином и миозином нарушаются, акто-миозиновый комплекс распадается, гладкий миоцит расслабляется.
Гладкие миоциты синтезируют протеогликаны, гликопротеиды, проколлаген, проэластин, из которых формируются коллагеновые и эластические волокна и основное вещество межклеточного матрикса.
Взаимодействие миоцитов осуществляется с помощью цитоплазматических мостиков, взаимных впячиваний, нексусов, десмосом или простых участков мембранных контактов клеточных поверхностей.
Регенерация гладкой мышечной ткани
Гладкая мышечная ткань висцерального и сосудистого видов обладает значительной чувствительностью к воздействию экстремальных факторов.
В активированных миоцитах возрастает уровень биосинтетических процессов, морфологическим выражением которых являются синтез сократительных белков, укрупнение и гиперхроматоз ядра, гипертрофия ядрышка, возрастание показателей ядерно-цитоплазменного отношения, увеличение количества свободных рибосом и полисом, активация ферментов, аэробного и анаэробного фосфорилирования, мембранного транспорта. Клеточная регенерация осуществляется как за счет дифференцированных клеток, обладающих способностью вступать в митотический цикл, так и за счет активизации камбиальных элементов (миоцитов малого объема).
При действии ряда повреждающих факторов отмечается фенотипическая трансформация контрактильных миоцитов в секреторные. Данная трансформация часто наблюдается при повреждении интимы сосудов, формировании ее гиперплазии при развитии атеросклероза.
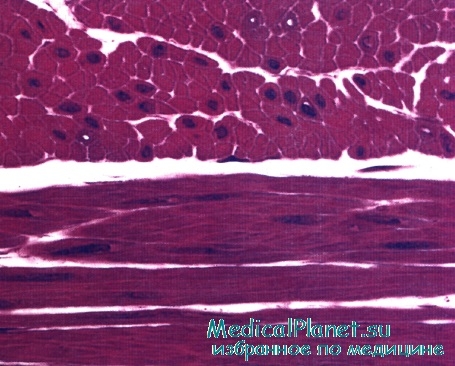
Гладкая мышечная ткань в поперечном (наверху) и продольном (внизу) разрезах. Обратите внимание на центрально расположенные ядра. Во многих клетках ядра не попали в срез.
Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
ЛЕКЦИЯ 8. МЫШЕЧНЫЕ ТКАНИ
Мышечные ткани классифицируются на гладкую и исчерченную, или поперечнополосатую. Поперечнополосатая подразделяется на скелетную и сердечную. В зависимости от происхождения мышечные ткани делятся на 5 типов: 1) мезенхимные (гладкая мышечная ткань); 2) эпидермал-ные (гладкая мышечная ткань);3) нейральные (гладкая мышечная ткань); 4)целомические (сердечная); 5) соматические или миотомные (скелетная поперечнополосатая).
ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ, РАЗВИВАЮЩАЯСЯ ИЗ СПЛАНХНОТОМНОЙ МЕЗЕНХИМЫ локализуется в стенках полых органов (желудка, кровеносных сосудах, дыхательных путях и др.) и неполых органах (в мышце реснич-ного тела глаза млекопитающих). Клетки гладкой мышечной ткани РАЗВИВАЮТСЯ из мезенхимоцитов, которые утрачивают отростки. В них развиваются комплекс Гольджи, митохондрии, гранулярная ЭПС и миофил-менты. В это время на гранулярной ЭПС активно синтезируется коллаген V типа, за счет которого вокруг клетки формируется базальная мембрана. При дальнейшей дифференцировке органеллы общего значения атрофируются, снижается синтез молекул коллагена в клетке, но повышается синтез сократительных белков миофиламентов. СТРОЕНИЕ ГЛАДКОЙ МЫШЕЧНОЙ ТКАНИ. Она состоит из гладких миоцитов, имеющих веретеновидную форму, длиной от 20 до 500 мкм. диаметром 6-8 мкм. Снаружи миоциты покрыты плазмолеммой и базальной мембраной. Моциты плотно прилежат друг к другу. Между ними имеются контакты — нексусы. В том месте, где имеются нексусы, в базальной мембране оболочки миоцитов есть отверстия. В этом месте плазмолемма одного миоцита приближается к плазмолемме другого миоцита на расстояние 2-3 нм. Через нексусы происходит обмен ионов, транспорт молекул воды, передача сократительного импульса.
Читайте также: Ткань для штор бархат синий
Снаружи миоциты покрыты коллагеном V типа, образующим экзоцитоскелет клетки. Цитоплазма миоцитов окрашивается оксифильно. В ней содержатся слабо развитые органеллы общего значения: гранулярная ЭПС, комплекс Гольджи, гладкая ЭПС, клеточный центр, лизосомы. Эти органеллы раполагаются у полюсов ядра. Хорошо развитые органеллы — митохондрии. ЯДРА имеют палочковидную форму. В миоцитах хорошо развиты миофиламенты, являющиеся сократительным аппаратом клеток. Среди миофиламентов имеются 1) тонкие, актиновые, состоящие из белка актина; 2) толстые миозиновые, состоящие из сократительного белка миозина, которые появляются только после поступления к клетке импульса; 3) промежуточные филаменты, состоящие из коннектина и небулина. В миоцитах отсутствует исчерченность потому, что все вышеперечисленные филаменты расположены неупорядоченно. АКТИНОВЫЕ ФИЛАМЕНТЫ соединяются друг с другом и с плазмолеммой при помощи плотных телец. В тех местах, где они соединяются друг с другом, в тельцах содержится альфа-актинин; в тех местах, где филаменты соединяются с плазмолеммой — в тельцах содержится винкулин. Расположение актиновых филаментов преимущественно продольное, но они могут располагаться под углом по отношению к продольной оси. Миозиновые филаменты тоже располагаются преимущественно продольно. Филаменты располагаются так, что концы актиновых располагаются между концами миозиновых филаментов. ФУНКЦИЯ ФИЛАМЕНТОВ — сократительная. Процесс сокращения осуществляется следующим образом: после поступления сократительного импульса пиноцитозные пузырьки, содержащие ионы кальция, приближаются к филаментам; ионы кальция запускают сократительный процесс, который заключается в том, что концы актиновых филаментов продвигаются глубже между концами миозиновых филаментов. Сила тяги прилагается к плазмолемме, с которой актиновые филаменты связаны при помощи плотных телец, в результате этого миоцит сокращается. ФУНКЦИИ МИОЦИТОВ: 1) сократительная (способность к длительному сокращению); 2) секреторная (секретируют коллаген V типа, эластин, протеогликаны, так как имеют гранулярную ЭПС).
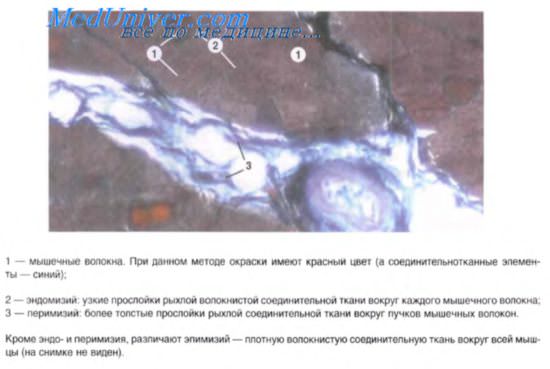
РЕГЕНЕРАЦИЯ гладкой мышечной ткани осуществляется 2 путями: 1) митотическое деление миоцитов; 2) преобразование в гладкие миоциты миофибробластов. СТРОЕНИЕ ГЛАДКОЙ МЫШЕЧНОЙ ТКАНИ КАК ОРГАНА. В стенке полых органов гладкие миоциты образуют пучки. Эти пучки окружены прослойками рыхлой соединительной ткани, которая называется перимизием. Прослойка соединительной ткани вокруг всего пласта мышечной ткани называется эпимизием. В перимизии и эпимизии проходят кровеносные и лимфатические сосуды и нервные волокна. ИННЕРВАЦИЯ ГЛАДКОЙ МЫШЕЧНОЙ ТКАНИ осществляется вегетативной нервной системой, поэтому сокращения гладкой мускулатуры не подчиняются воле человека (непроизвольные). К гладкой мышечной ткани подходят чувствительные (афферентные) и двигательные (эфферентные) нервные волокна. Эфферентные нервные волокна заканчиваются двигательными нервными окончаниями в прослойке соединительной ткани. При поступлении импульса из окончаний выделяются медиаторы, которые, диффузно распространяясь, достигают миоцитов, вызывая их сокращение. ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ ЭПИДЕРМАЛЬНОГО ПРОИСХОЖДЕНИЯ находится в концевых отделах и мелких протоках желез, которые развиваются из кожной эктодермы (слюнные, потовые, молочные и слезные железы). Гладкие миоциты (миоэпителиоциты) располагаются между базальной поверхностью железистых клеток и базальной мембраной, охватывая базальную часть гландулоцитов своими отростками. При сокращении этих отростков сдавливается базальная часть гландулоцитов, благодаря чему из железистых клеток выделяется секрет. ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ НЕЙРАЛЬНОГО ПРОИСХОЖДЕНИЯ развивается из глазных бокалов, вырастающих из нервной трубки. Эта мышечная ткань образует всего 2 мышцы, расположенные в радужной оболочке глаза: мышцу суживающую зрачок и мышцу расширяющую зрачок. Существует мнение, что мышцы радужки развиваются из нейроглии. ПОПЕРЕЧНОПОЛОСАТАЯ СКЕЛЕТНАЯ МЫШЕЧНАЯ ТКАНЬ развивается из миотомов мезодермальных сомитов, поэтому называется соматической. Клетки миотомов дифференцируются в двух направлениях: 1) из одних образуются миосателлитоциты; 2) из других образуются миосимпласты. ОБРАЗОВАНИЕ МИОСИМПЛАСТОВ. Клетки миотомов дифференцируются в миобласты, которые сливаются вместе и образуются мышечные трубочки. В процессе созревания мышечные трубочки превращаются в миосимпласты. При этом ядра смещаются к периферии, а миофибриллы — к центру. СТРОЕНИЕ МЫШЕЧНОГО ВОЛОКНА. Мышечное волокно (miofibra) состоит из 2 компонентов: 1) миосателлитоцитов и 2) миосимпласта. Мышечное волокно имеет примерно такую же длину, как и сама мышца, диаметр — 20-50 мкм. Волокно снаружи покрыто оболочкой — сарколеммой, состоящей из 2 мембран. Наружная мамбрана называется базальной мембраной, а внутренняя — плазмолеммой. Между этими двумя мембранами располагаются миосателлитоциты. ЯДРА МЫШЕЧНЫХ ВОЛОКОН располагаются под плазмолеммой, их количество может достигать нескольких десятков тысяч. Имеют вытянутую форму, не обладают способностью к дальнейшему митотическому делению. ЦИТОПЛАЗМА мышечного волокна называется САРКОПЛАЗМОЙ. В саркоплазме содержится большое количество миоглобина, включений гликогена и липидов; имеются органеллы общего значения, одни из которых развиты хорошо, другие — хуже. Такие органеллы как комплекс Гольджи, гранулярная ЭПС, лизосомы развиты слабо и располагаются у полюсов ядер. Хорошо развиты митохондрии и гладкая ЭПС. В мышечных волокнах хорошо развиты миофибриллы, являющиеся сократительным аппаратом волокна. В миофибриллах имеется исчерченность потому, что миофиламенты в них расположены в строго определенном порядке (в отличии от гладкой мускулатуры). В миофибриллах 2 вида миофиламентов: 1) тонкие актиновые, состоят из белка актина, тропонина и тропомиозина; 2) толстые миозиновые состоят из белка миозина. Актиновые филаменты располагаются продольно, их концы находятся на одинаковом уровне и несколько заходят между концами миозиновых филаментов. Вокруг каждого миозинового филамента расположено 6 концов актиновых филаментов. В мышечном волокне имеется цитоскелет, включающий промежуточные нити (филаменты), телофрагму, мезофрагму, сарколемму. Благодаря цитоскелету одинаковые структуры миофибрилл (актиновые, миозиновые филаменты и др.) располагаются упорядоченно. Тот участок миофибриллы, в котором находятся только актиновые филаменты, называется диском I (изотропный или светлый диск). Через центр диска I проходит Z-полоска, или телофрагма толщиной около 100 нм и состоящая из альфа-актинина. К телофрагме прикрепляются актиновые нити (зона прикрепления тонких нитей). Миозиновые филаменты тоже располагаются в строго определенном порядке. Их концы также находятся на одном уровне. Миозиновые филаменты вместе с заходящими между ними концами актиновых филаментов образуют диск А (анизотропный диск, обладающий двулучепреломлением). Диск А также разделяется мезофрагмой, аналогичной телофрагме и состоящей из М-белка (миомизина). В средней части диска А имеется Н-полоска, ограниченная концами актиновых филаментов, заходящих между концами миозиновых нитей. Поэтому чем ближе концы актиновых филаментов расположены друг к другу, тем уже Н-полоска. САРКОМЕР — это структурная и функциональная единица миофибрилл, представляющая собой участок, расположенный между двумя телофрагмами. Формула саркомера: 1,5 диса I + диск А + 1,5 диска I. Миофибриллы окружены хорошо развитыми митохондриями и хорошо развитой гладкой ЭПС. ГЛАДКАЯ ЭПС образует систему L-канальцев, образующих в каждом диске сложные структуры. Эти структуры состоят из L-канальцев расположенных вдоль миофибрил и соединяющихся с поперечно направленными L-канальцами (латеральными цистернами). ФУНКЦИИ гладкой ЭПС(системы L-канальцев): 1) транспортная; 2) синтез липидов и гликогена; 3) депонирование ионов кальция. Т-КАНАЛЫ — это впячивания плазмолеммы. На границе дисков из плазмолеммы вглубь волокна происходит впячивание в виде трубочки, располагающейся между двумя латеральными цистернами. ТРИАДА включает: 1) Т-канал и 2) 2 латеральные цистерны гладкой ЭПС. ФУНКЦИЯ ТРИАД заключается в том, что в расслабленном состоянии миофибрил в латеральных цистернах накапливаются ионы кальция; в тот момент, когда по плазмолемме движется импульс (потенциал действия), он переходит на Т-каналы. При движении импульса по Т-каналу из латеральных цистерн выходят ионы кальция. Без ионов кальция невозможно сокращение миофибрил, потому что в актиновых филаментах центры взаимодействия с миозиновыми нитями заблокированы тропомиозином. Ионы кальция осуществляют разблокирование этих центров, после чего начинается взаимодействие актиновых нитей с миозиновыми и начинается сокращение. МЕХАНИЗМ СОКРАЩЕНИЯ МИОФИБРИЛЛ. При взаимодействии актиновых филаментов с миозиновыми происходит разблокирование ионами Са центров сцепления актиновых филаментов с головками молекул миозина, после чего эти выросты присоединяются к центрам сцепления на актиновых нитях и как веслом осуществляют движение актиновых филаментов между концами миозиновых. В это время телофрагма приближается к концам миозиновых филаментов, поскольку концы актиновых филаментов тоже приближаются к мезофрагме и друг к другу, постольку происходит сужение Н-полоски. Таким образом, во время сокращения миофибрилл происходит сужение диска I и Н-полоски. После прекращения потенциала действия ионы кальция возвращаются в L-канальцы гладкой ЭПС, тропомиозин снова блокирует в актиновых филаментах центры взаимодействия с миозиновыми нитями. Это приводит к прекращению сокращения миофибрил, происходит их расслабление, т.е. актиновые нити возвращаются в исходное положение, восстанавливается ширина диска I и Н-полоски. МИОСАТЕЛЛИТОЦИТЫ мышечного волокна располагаются между базальной мембраной и плазмолеммой сарколеммы. Эти клетки имеют овальную форму, их овальное ядро окружено тонким слоем бедной органеллами и слабо окрашиваемой цитоплазмы. ФУНКЦИЯ миосателлитоцитов — это камбиальные клетки, участвующие в регенерации мышечных волокон при их повреждении. СТРОЕНИЕ МЫШЦЫ КАК ОРГАНА. Каждая мышца тела человека представляет собой своеобразный орган, имеющий свою структуру. Каждая мышца состоит из мышечных волокон. Каждое волокно окружено тонкой прослойкой рыхлой соединительной ткани — эндомизием. В эндомизии проходят кровеносные и лимфатические сосуды и нервные волокна. Мышечное волокно вместе с сосудами и нервными волокнами называется «мион». Несколько мышечных волкон образуют пучок, окруженный слоем рыхлой соединительной ткани, называемой перимизием. Вся мышца окружена прослойкой соединительной ткани, называемой эпимизием. СВЯЗЬ МЫШЕЧНЫХ ВОЛОКОН С КОЛЛАГЕНОВЫМИ ВОЛОКНАМИ СУХОЖИЛИЙ. На концах мышечных волокон имеются впячивания сарколеммы. В эти впячивания входят коллагеновые и ретикулярные волокна сухожилий. Ретикулярные волокна прободают базальную мембрану и при помощи молекулярных сцеплений соединяются с плазмолеммой. Затем эти волокна возвращаются в просвет впячивания и оплетают коллагеновые волокна сухожилия, как бы привязывая их к мышечному волокну. Коллагеновые волокна образуют сухожилия, которые прикрепляются к костному скелету. ТИПЫ МЫШЕЧНЫХ ВОЛОКОН. Имееися 2 основных типа мышечных волокон: 1) I тип (красные волокна) и II тип (белые волокна). Они различаются главным образом по быстроте сокращения, содержанию миоглобина, гликогена и активности ферментов. 1-й ТИП (красные волокна) характеризуются большим содержанием миоглобина (поэтому они красные), высокой активностью сукцинатдегидрогеназы, АТФ-зой медленного типа, не столь богатым содержанием гликогена, длительностью сокращения и малой утомляемостью. 2-й ТИП (белые волокна) отличаются малым содержанием миоглобина, низкой активностью сукцинатдегидрогеназы, АТФ-зой быстрого типа, богатым содержанием гликогена, быстрым сокращением и большой утомляемостью. Медленный (красный) и быстрый (белый) тип мышечных волокон иннервируются разными типами моторных нейронов: медленным и быстрым. Кроме 1-го и 2-го типов мышечных волокон имеются промежуточные, обладающие совйствами тех и других. В каждой мышце имеются все типы мышечных волокон. Их количество может меняться и зависит от физической нагрузки. РЕГЕНЕРАЦИЯ ПОПЕРЕЧНОПОЛОСАТОЙ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ. При повреждении (разрыве) мышечных волокон их концы на месте повреждения подвергаются некрозу. После разрыва к обрывкам волокон поступают макрофаги, которые фагцитируют некротизированные участки, очищая их от мертвой ткани. После этого процесс регенерации осуществляется 2 путями: 1) за счет повышения реактивности в мышечных волокнах и образования мышечных почек в местах разрыва; 2) за счет миосателлитоцитов. 1-й ПУТЬ характеризуется тем, что на концах разорванных волокон гипертрофируется гранулярная ЭПС, на поверхности которой синтезируются белки миофибрил, мембранных структур внутри волокна и сарколеммы. В результате этого концы мышечных волокон утолщаются и преобразуются в мышечные почки. Эти почки по мере их увеличения приближаются друг к другу от одного оборванного конца к другому, наконец почки соединяются и срастаются. Между тем за счет клеток эндомизия происходит новообразование соединительной ткани между растущими навстречу друг к другу мышечными почками. Поэтому к моменту соединения мышечных почек формируется соединительнотканная прослойка, которая войдет в состав мышечного волокна. Следовательно, формируется соединительнотканный рубец. 2-й ПУТЬ регенерации заключается в том, что миосателлитоциты покидают места своего обитания и подвергаются дифференцировке, в результате которой превращаются в миобласты. Часть миобластов присоединяется к мышечным почкам, часть соединяется в мышечные трубочки, которые дифференцируются в новые мышечные волокна. Таким образом, при репаративной регенерации мышц восстанавливаются старые мышечные волокна и образуются новые. ИННЕРВАЦИЯ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ осуществляется двигательными и чувствительными нервными волокнами, заканчивающимися нервными окончаниями.ДВИГАТЕЛЬНЫЕ (моторные) нервные окончания являются концевыми приборами аксонов мотроных нервных клеток передних рогов спинного мозга. Конец аксона, подходя к мышечному волокну делится на несколько веточек (терминалей). Терминали прободают базальную мембрану сарколеммы и далее погружаются вглубь мышечного волокна, увлекая за собой плазмолемму. В результате этого образуется нервномышечное окончание (моторная бляшка). СТРОЕНИЕ НЕРВНОМЫШЕЧНОГО окончания. В нервномышечном окончании имется две части (полюса): нервная и мышечная. Между нервной и мышечной частями имеется синаптическая щель. В нервной части (терминалях аксона моторного нейрона) имеются митохондрии и синаптические пузырьки, заполненные медиатором-ацетилхолином. В мышечной части нервномышечного окончания есть митохондрии, скопление ядер, отсутствуют миофибриллы. Синаптическая щель шириной 50 нм ограничена пресинаптической мембраной (плазмолеммой аксона) и постсинаптической мембраной (плазмолеммой мышечного волокна). Постсинаптическая мембрана образует складки (вторичные синаптические щели), на ней имеются рецепторы к ацетилхолину и фермент — ацетилхолинэстераза. ФУНКЦИЯ нервномышечных окончаний. Импульс движется по плазмо — лемме аксона (пресинаптической мембране). В это время синаптические пузырьки с ацетилхолином подходят к плазмолемме, из пузырьков ацетилхолин изливается в сиаптическую щель и захватывается рецепторам постсинаптической мембраны. Это повышает проницаемость этой мембраны (плазмолеммы мышечного волокна), в результате этого ионы натрия с наружной поверхности плазмолеммы переходят на внутреннюю, а ионы калия переходят на наружную поверхность — это и есть олна деполяризации или нервный импульс (потенциал действия). После возникновения потенциала действия ацетилхолинэстераза постсинаптической мембраны разрушает ацетилхолин и прекращается переход импульса через синаптическую щель. ЧУВСТВИТЕЛЬНЫМИ НЕРВНЫМИ ОКОНЧАНИЯМИ (нервно-мышечными веретенами-fusi neuro-muscularis) заканчиваются дендриты чувствительных нейронов спинномозговых узлов. Нервно-мышечные веретена покрыты со единительнотканной капсулой, внутри которой имеются 2 типа интрафузальных (внутриверетенных) мышечных волокон: 1) с ядерной сумкой (в центе волокна утолщение, в котором имеется скопление ядер), они более длинные и более толстые; 2) с ядерной цепочкой (ядра ввиде цепочки располагаются по центру волокна), они тонше и короче. В окончания проникают толстые нервные волокна, которые кольце образно оплетают оба вида интрафузальных мышечных волокон и тонкие нервные волокна, заканчивающиеся гроздъевидными окончаниями на мышечных волокнах с ядерной цепочкой. На концах интрафузальных волокон имеются миофибриллы и к ним подходят двигательные нервные окончания. Сокращения интрафузальных волокон не обладают большой силой и не суммируются с остальными (экстрафузальными) волокнами мышцы. ФУНКЦИЯ нервно-мышечных веретен заключается в восприятии скорости и силы растяжения мышцы. Если сила растяжения такова, что угрожает разрывом мышцы, то на сокращающиеся мышцы-антогонисты от этих окончаний рефлекторно поступают тормозные импульсы. СЕРДЕЧНАЯ МЫШЕЧНАЯ ТКАНЬ развивается из переднего отдела висцеральных листков спланхнотома. Из этих листков выделяются 2 миоэпикардиальных пластинки: правая и левая. Клетки миоэпикардиальных пластинок ифференцируются в двух направлениях: из одних развивается мезотелий, покрывающий эпикард, из других — кардиомиоциты пяти разновидностей; 1 — сократительные, 2 — пейсмекерные, 3 — проводящие, 4 — промежуточные, 5 — секреторные, или эндокринные.
Читайте также: Обмен газа в тканях это
СТРОЕНИЕ КАРДИОМИОЦИТОВ. Кардиомиоциты имеют цилиндрическую форму, длиной 50-120 мкм, диаметром 10-20 мкм. Кардиомиоциты соеди няются концами друг с другом и образуют функциональные сердечные мышечные волокна. Места соединения кардиомиоцитов называются вставочными дисками. В дисках имеются интердигитации, десмосомы, места прикрепления актиновых филаментов и нексусы. Через нексусы происходит обмен веществ между кардиомиоцитами. Снаружи кардиомиоциты покрыты сарколеммой, состоящей из наружной (базальной) мембраны и плазмолеммы. От боковых поверхностей кардиомиоцитов отходят отростки, вплетающиеся в боковые поверхности кардиомиоцитов соседнего волокна. Это мышечные анастомозы. ЯДРА кардиомиоцитов (одно-два), овальной формы, обычно полиплоидные, располагаются в центре клетки. МИОФИБРИЛЛЫ локализованы по периферии. ОРГАНЕЛЛЫ — одни развиты слабо (гранулярная ЭПС, комплекс Гольджи, лизосомы), другие — хорошо (митохондрии, гладкая ЭПС, миофибриллы). В оксифильной ЦИТОПЛАЗМЕ имеются включения миоглобина, гликогена и липидов.
СТРОЕНИЕ МИОФИБРИЛЛ такое же как и в скелетной мышечной ткани. Актиновые филаменты формируют светлый диск ( I ), разделенный телофрагмой, за счет миозиновых филаментов и концов актиновых образуется диск А (анизотропный), разделенный мезофрагмой. В средней части диска А имеется Н-полоска, ограниченная концами актиновых филаментов. Волокна сердечной мышцы отличаются от волокон скелетной мускулатуры тем, что состоят они из отдельных клеток-кардиомиоцитов, наличием мышечных анастомозов, центральным раположением ядер (в волокне скелетной мышцы — под сарколеммой), увеличенной толщиной диаметра Т-каналов, так как в их состав входит и плазмолемма и базальная мембрана (в волокнах скелетной мышцы — только плазмолемма). ПРОЦЕСС СОКРАЩЕНИЯ в волокнах сердечной мышцы осуществляется по такому же принципу, как и волокнах скелетной мышечной ткани.
ПРОВОДЯЩИЕ КАРДИОМИОЦИТЫ характеризуются более толстым диаметром (до 50 мкм), более светлой цитоплазмой, центральным или эксцентричным расположением ядер, малым содержанием миофибрилл, более простым устройством вставочных дисков. В дисках меньше десмосом, интердигитаций, нексусов и мест прикрепления актиновых филаментов. В проводящих кардиомиоцитах отсутствуют Т-каналы. Проводящие кардиоиоциты могут соединяться друг с другом не только своими концами, но и боковыми поверхностями. ФУНКЦИЯ проводящих кардиомиоцитов заключается в выработке и передаче сократительного импульса на сократительные кардиомиоциты. ЭНДОКРИННЫЕ КАРДИОМИОЦИТЫ располагаются только в предсердиях, имеют более отростчатую форму, слабо развитые миофибриллы, вставочные диски, Т-каналы. В них хорошо развиты гранулярная ЭПС, комплекс Гольджи и митохондрии, в их цитоплазме имеются гранулы секрета. ФУНКЦИЯ эндокринных кардиомиоцитов — секреция предсердного натрийуретического фактора (ПНФ), который регулирует сократимость сердечной мышцы, объем циркулирующей жидкости, артериальное давление, диурез. РЕГЕНЕРАЦИЯ сердечной мышечной ткани только физиологическая, внутриклеточная. При повреждении волокон сердечной мышцы, они не восстанавливаются, а замещаются соединительной тканью (гистотипическая регенерация).