
Особенности анатомии и морфологии листа как органа фотосинтеза
В зависимости от вида растений и условий их произрастания листья отличаются большим разнообразием. Однако можно выделить общие анатомические особенности, обеспечивающие возможность эффективного фотосинтеза.
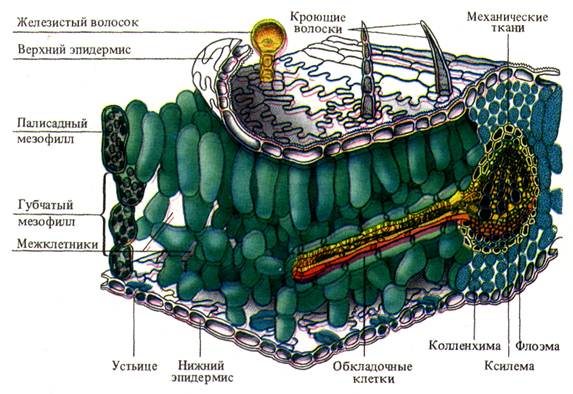
1. Наличие покровной ткани — эпидермиса, защищающего лист от излишней потери воды. Клетки нижнего и верхнего эпидермиса лишены хлоропластов, имеют крупные вакуоли, которые, подобно линзам, фокусируют свет на расположенную глубже хлорофиллоносную ткань. Деятельность клеток мезофилла основана не только на их хорошем освещении, она зависит также от поступления СО2. Нижний эпидермис, реже и верхний, имеют большое количество устьиц. Щели открытых устьиц занимают примерно 1 % площади листовой пластинки, диффузия СО2 внутрь листа идет через них сравнительно быстро. Отдельное устьице позволяет за 1 с поступить в лист 2500 млрд. молекул СО2. Поверхность листа поглощает СО2 только в 1,5-2 раза меньше, чем открытая поверхность щелей той же площади, хотя открытые устьица составляют лишь сотую часть поверхности. Такая высокая скорость связана с особенностями диффузии газов через мелкие отверстия, находящиеся на значительном расстоянии друг от друга, за счет краевого эффекта.
2. Наличие специализированной фотосинтетической ткани — хлоренхимы. Основная хлорофиллоносная ткань — палисадная паренхима — расположена обычно на освещаемой части листа. Вытянутость клеток и перпендикулярное расположение их к эпидермису обеспечивают увеличение поверхности, вдоль которой могут располагаться хлоропласты, не затеняя друг друга, а также облегчают отток ассимилятов. В каждой клетке полисадной паренхимы находится 30-40 хлоропластов. Губчатая ткань характеризуется меньшим содержанием хлоропластов (примерно 20 на клетку) и сильно развитой системой межклетников. Объем межклетников составляет 15-20 % общего объема листа и образует внутреннюю газовую среду, которая при помощи устьичных щелей сообщается с атмосферой. За счет межклетников значительно возрастает внутренняя рабочая поверхность, через которую каждой клеткой паренхимы поглощается СО2. Она в 8-12 раз больше, чем наружная поверхность листа. Хлоропласты как основные светоулавливающие органеллы еще в большей степени увеличивают светопоглощающую поверхность листа. На 1 см 2 листа приходится примерно 200 см 2 поверхности хлоропластов. Это является выражением общебиологической особенности организации — создание больших внутренних рабочих поверхностей при сравнительно малых наружных испаряющих площадях за счет, как было отмечено, затрат небольших количеств материала.
3. Наличие сильно развитой густой системы жилок — проводящих путей, что обеспечивает быстрый отток ассимилятов и снабжение фотосинтезирующих клеток водой и необходимыми минеральными веществами.
В зависимости от внешних условий, при которых происходят формирование и функционирование листьев, анатомическое строение их может существенно различаться. Листья, формирующиеся в условиях недостаточной влагообеспеченности, имеют ксероморфную структуру. В зависимости от освещения меняется соотношение между полисадной и губчатой паренхимой в мезофилле. Имеются и другие приспособления для функционирования листа в определенных условиях. Еще более существенные отклонения от типичного строения листа связаны с физиолого-биохимическими особенностями фиксации СО2 у С4-растений, к которым относятся кукуруза, сахарный тростник, ряд злостных сорняков наших полей.
Организация фотосинтетического аппарата на уровне листа может быть также охарактеризована на основе анализа его мезоструктуры. Понятие «мезоструктура» предложено А.Т. Мокроносовым в 1975 г. Оно охватывает целый ряд морфофизиологиических характеристик листа, позволяющих оценить ассимиляционную способность листа в целом. Основными показателями мезоструктуры листа являются: площадь листа, число клеток хлоренхимы на единицу площади листа, число хлоропластов в клетке и их объем, площадь поверхности хлоропластов, а также содержание хлорофилла в расчете на единицу площади листа, содержание ферментов углеродного цикла фотосинтеза в листе и их активность, общая интенсивность фотосинтеза. Показатели мезоструктуры листа могут значительно варьировать в зависимости от внешних факторов среды, а также от физиологического состояния растений.
Лист представляет собой уникальную оптическую систему — ловушку для света. Как видно из рисунка, лист поглощает значительную часть излучения в видимой области спектра (400-700 нм). Большую часть синего и красного света поглощают пигменты хлоропластов первых слоев клеток хлоренхимы. Свет, не поглощенный в верхних слоях листа, обогащен лучами зеленой области спектра. Его многократное отражение от стенок мезофильных клеток в толще листа значительно увеличивает оптический путь и повышает вероятность поглощения лучей этой области спектра, несмотря на то что коэффициент поглощения хлорофилла в зеленой области значительно меньше, чем в синей и красной.
Читайте также: Чем отмыть ткань микрофибру
Структура листа обеспечивает поступление углекислого газа из воздуха для реакций фотосинтетической ассимиляции углерода. Газообмен мезофилла листа с окружающим воздухом осуществляется через устьица. Количество устьиц, их свойства (величина устьичной щели, устьичное сопротивление и др.) являются важными факторами регуляции фотосинтеза. Значительный объем подустьичного пространства, а также достаточно рыхлое расположение мезофильных клеток в листе и существование больших межклетников в ткани хлоренхимы позволяют создать большие воздушные запасы в толще листа для непрерывного снабжения углекислым газом углеродных циклов фотосинтеза.
Лист — донор ассимилятов в растении. Отток органических веществ, образовавшихся в процессе фотосинтеза, происходит по клеткам сосудистых пучков флоэмы. Транспорт ассимилятов из автотрофных клеток предполагает вынос сахарозы (основного транспортного соединения фотосинтеза) из авто-трофной клетки в апопласт и последующую загрузку флоэмы листа.
Структурная организация фотосинтетического аппарата, строение листа как органа фотосинтеза, ультраструктура и онтогенез хлоропластов
Ответы на вопросы по физиологии растений
Основные принципы действия регуляторных механизмов растительной клетки; компартментация каталитических систем и метаболических фондов протопласта как один из механизмов регуляции клеточного метаболизма.
Клетка – это мельчайшая единица жизни, характеризующаяся определенным типом обмена веществ, самостоятельным энергетическим циклом и способностью к саморегуляции и саморазвитию на основе взаимодействия различных своих компо-нентов и взаимодействия с внешними условиями. В живой клетке нет изолированных процессов. Основой этого взаимодействия служит непрерывный обмен продуктами реакций, образующимися в органоидах определенного вида и поступающими в цитоплазму и другие органоиды.
Сложное строение растительной клетки, дифференцированной на большое количество специализированных органоидов и структур в соответствии с выполняемыми ее функциями, требует и совершенных систем регуляции. Регуляция обеспечивает целостность и гомеостаз организма, т. е. сохранение постоянства параметров внутренней среды, а также создает условия для его развития (эпигенеза).
В ходе эволюции сначала возникли внутриклеточные системы регуляции. К ним относятся регуляция на уровне ферментов, генетическая и мембранная регуляции. Все эти системы регуляции тесно связаны между собой. Принципы действия регуляторных механизмов клетки:
– принцип взаимодействия компонентов цитоплазмы.
В основе всех форм внутриклеточной регуляции лежит единый первичный принцип – рецепторно-конформационный принцип. Белковая молекула – фермент, рецептор или регуляторный белок — «узнает» специфический для нее фактор и, взаимодействуя с ним, изменяет свою конфигурацию. В мультикомпонентных комплексах ферментов, генов и мембран конформационные изменения молекул-рецепторов кооперативно передаются на весь комплекс, влияя на его функциональную активность.
Важнейшими в регуляторных механизмах клетки является принцип компар-тментации, обуславливающий высокую степень временной и пространственной ор-ганизованности метаболизма, и взаимодействия компонентов цитоплазмы. Взаимо-действие обеспечивается наличием гиалоплазмы и эндоплазматической сети.
Компартменты – дифференцированные, специализированные участки, или отсеки в протоплазме, различающиеся по степени активности содержащихся в них химических соединений и систем, регулирующих их превращения. Явление компартментации обуславливают клеточные мембраны, которые выполняют функцию расчленения биохимических процессов, разделения их между различными компонентами протоплазмы и пространственного размещения в объеме клетки фондов метаболитов и ферментов.
В растительной клетке имеется три основных компартмента: свободное пространство (СП), цитоплазма и вакуоль. Компартментальны отдельные участки клетки и органелл, отдельные участки мембран, отдельные фонды метаболитов, ферментов и всех других морфологических и химических компонентов протопласта.
Характерная особенность компартментированных метаболитов и ферментов — их подвижность, что достигается включением метаболитов в везикулы и мелкие вакуоли и концентрацией их в трубочках эндоплазматического ретикулума и других трубчатых структурах. Это способствует, например, отводу ассимилятов из хлоропластов без их соприкосновения со стромой цитоплазмы. Эффективность действия механизмов фотофосфорилирования в хлоропластах определяется компартментами, обеспечивающими образование протонного градиента. Компартментальны в данном случае мембранные диски гран хлоропластов. Практически все основные пути метаболизма углеводов прочно ассоциированы с определенными участками плазматических мембран. В клетке существуют компартменты, содержащие системы гликолиза, цикла Кребса и пентозофосфатного шунта. Так все участки цикла Кребса локализованы в матриксе или во внутренней мембране митохондрий. Аналогичные закономерности установлены для процессов биосинтеза и превращения аминокислот, поглощения и выделения минеральных веществ, воды, регуляции действия имеющихся в клетке ферментов и т. п.
Читайте также: Соединение двух кусков ткани незаметно
С появлением многоклеточных организмов развиваются и совершенствуются межклеточные системы регуляции. Они включают в себя по крайней мере трофическую, гормональную и электрофизиологическую системы. Такие взаимодействия сразу обнаруживаются при культивировании частей и органов растений в изолированном виде. Для поддержания жизни изолированных частей в инкубационную среду необходимо добавлять трофические и гормональные факторы, в норме поступающие из других органов целого растения.
Важным также является взаимная координация процессов, реакций, координация накопления и выделения веществ и энергии клеткой. Так, нуклеиновые кислоты играют роль хранителя и передатчика наследственной информации (ДНК) и посредника ее реализации (РНК). Белки несут функции рабочих механизмов – это ферменты, которые входят в состав клеточных структур и многих активных соединений. Основной механизм регуляции внутриклеточных процессов связан с влиянием на ферменты разнообразных факторов. Это может осуществляться на генетическом уровне, если определяется состав ферментов или количество того или иного фермента в клетке. Второй тип регуляции – влияние на сам фермент, в результате чего активность его может стимулироваться или ингибироваться. Может иметь место также структурный уровень регуляции – влияние на образование и сборку клеточных структур (мембран, хлоропластов, рибосом, митохондрий и т. д.). Конкретными регуляторами внутриклеточных процессов могут быть фитогормоны, которые вырабатываются клетками или окружающими их другими клетками, или продукты реакций по принципу обратной связи (аллостерический эффект). Регуляция может осуществляться также путем транспортировки в клетку продуктов метаболизма и их предшественников, ионов, изменения формы регулируемого фермента (конформаций).
Структурная организация фотосинтетического аппарата, строение листа как органа фотосинтеза, ультраструктура и онтогенез хлоропластов
«Зеленый лист, или, вернее, микроскопическое зеленое зерно хлорофилла является фокусом, точкой в мировом пространстве, в которую с одного конца притекает энергия солнца, а с другого берут начало все проявления жизни на земле. Растение — посредник между небом и землею. Оно истинный Прометей, похитивший огонь с неба» ( К. А. Тимирязев ).
Основой фотосинтетического аппарата служат:
− у высших зелёных растений внутриклеточные органеллы – хлоропласты;
− у большинства водорослей − специализированные внутриклеточные органеллы – хроматофоры;
− у фотосинтезирующих бактерий и синезелёных водорослей – тилакоиды (мембраны их содержат пигмент бактериохлорофилл или бактериовиридин, а также др. компоненты реакций фотосинтеза), погруженные в периферические слои цитоплазмы.
Хлоропласты занимают 20–30% объёма растительной клетки. У водорослей, например хламидомонады, имеется один хлоропласт, в клетке высших растений содержится от 10 до 70 хлоропластов и более.
Хлорофилл и каротиноиды, погруженные в тилакоидные мембраны хлоропластов, собраны в функциональные единицы – фотосистемы, каждая из которых содержит примерно 250 молекул пигментов. Фотосистема – совокупность ССК (светособирающего комплекса), фотохимического реакционного центра и переносчиков электрона.
ССК растений расположен в мембранах тилакоидов, у цианобактерий основная его часть вынесена за пределы мембран в прикреплённые к ним фикобилисомы — палочковидные полипептидно-пигментные комплексы, в которых находятся различные фикобилины: на периферии фикоэритрины, за ними фикоцианины и аллофикоцианины, последовательно передающие энергию на хлорофилл a реакционного центра.
Роль ССК состоит в том, чтобы собирать и передавать энергию квантов света на небольшое количество молекул (молекулы-ловушки) реакционных центров П680 и П700 которые и осуществляют фотохимические реакции.
Органом фотосинтеза у высших растений является лист. Кроме фотосинтеза в жизни растений лист выполняет функции газообмена, транспирации, синтеза ряда органических веществ, в том числе и фитогормонов.
Листья, будучи боковыми органами, как правило, имеют более или менее плоскую форму, что способствует созданию максимальной фотосинтезирующей поверхности.
Основной тканью листа является паренхимный комплекс клеток, несущий хлоропласты – мезофилл (греч. mesos – средний и филл) (рисунок 1). Остальные ткани листа обеспечивают работу мезофилла и поддерживают связь с окружающей средой. Покровная ткань (эпидермис) регулирует газообмен и транспирацию, защищает лист от внешних воздействий. Проводящие ткани осуществляют отток и приток веществ, поддерживают нормальное оводнение фотосинтезирующих клеток. Механические ткани совместно с живыми тургесцентными клетками мезофилла и эпидермиса образуют опорную систему листа.
Мезофилл. Представлен клетками основной паренхимы, расположенными между верхним и нижним эпидермисом (исключая механические ткани и проводящие пучки). Состоит из живых клеток, с тонкими оболочками, округлой или слегка вытянутой формы, реже с небольшими выростами. Благодаря многочисленным хлоропластам мезофилл окрашен в зеленый цвет, так называемая хлоренхима листа. Иногда оболочки клетки образуют складки внутрь полости клетки, что увеличивает рабочую поверхность хлоренхимных клеток.
Читайте также: Лимфоидная ткань слизистых оболочек кожи
У типичных дорсовентральных листьев мезофилл неоднороден и диффе-ренцирован на столбчатую (палисадную) и губчатую паренхиму.
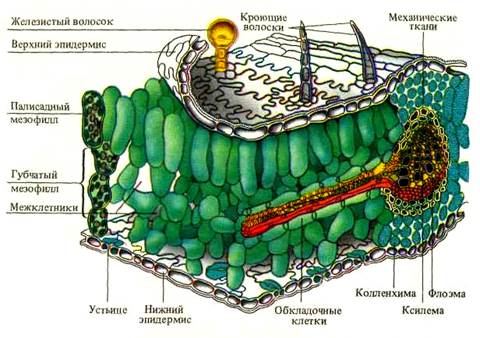
Рисунок 1 – схема строения листа
Клетки столбчатой паренхимы плотно сомкнуты, имеют продолговатую форму и располагаются перпендикулярно к верхней стороне листа. Столбчатый мезофилл может быть однослойный, иногда двуслойный, редко (у светолюбивых растений) многослойный. Основная функция палисадной паренхимы – ассимиляция углекислого газа (фотосинтез). Палисадная ткань располагается в наилучших условиях освещения, непосредственно под верхней эпидермой. Благодаря тому, что ее клетки вытянуты перпендикулярно к поверхности листа, у них имеется возможность регулировать направление и расположение хлоропластов таким образом, чтобы избежать повреждающего действия прямой солнечной радиации на фотосинтезирующий аппарат.
Губчатую паренхиму составляют относительно округлые клетки с большими межклетниками, которые занимают больший объем, чем сами клетки. Рыхлая структура мезофилла обусловливает большую общую площадь поверхности клеток, обращенной к межклетникам. Через крупные межклетники губчатого мезофилла идет газообмен.
Форма – овальные тельца длиной 5-10 мкм и диаметром 2-3 мкм. Количество в клетке листа – 15-20, у некоторых водорослей — 1-2 гигантских хлоропласта различной формы. Окраска – зеленая, обусловлена содержанием в хлоропластах хлорофилла.
Хлоропласты – двухмембранные органоиды (рисунок 2). Внутреннее содержимое хлоропластов − строма (матрикс) представляет собой гомогенную среду. Хлоропласты, как и другие органоиды, например, митохондрии, имеют ламеллярное строение, т.е. внутренние структуры органоида образованы инвагинациями внутренней мембраны – ламеллами. В хлоропластах внутренняя мембрана образует уплощенные инвагинации — тилакоиды, которые могут иметь форму дисков и в этом случае называются тилакоидами гран.
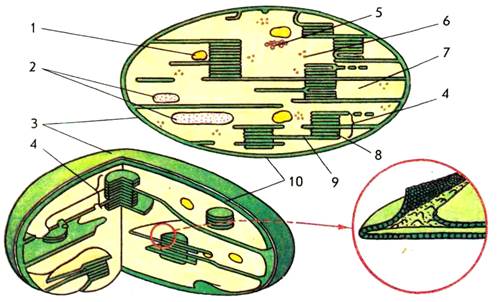
Рисунок 2 – схема строения хлоропласта:
1 – липидная капля; 2 – зерна крахмала; 3 – наружная мембрана; 4 – грана; 5 – ДНК; 6 – рибосома; 7 – строма; 8 – тилакоид граны; 9 – тилакоид стромы; 10 – внутренняя мембрана
Несколько лежащих друг над другом тилакоидов образуют стопку – грану. Другие тилакоиды, связывающие между собой граны или не контактирующие с ними, называются тилакоидами стромы. При таком строении значительно увеличивается фотоактивная поверхность хлоропластов и обеспечивается максимальное использование световой энергии. В мембранах тилакоидов локализованы зеленые (хлорофиллы), желтые и красные (каротиноиды) пигменты, компоненты редокс-цепей и запасания энергии, участвующие в поглощении и использовании энергии света.
Граны часто имеют форму цилиндров размером 2 мкм, в которых пигментно-липидные слои чередуются с белковыми слоями. Морфологически тилакоиды не гомогенны. На внутренней поверхности их мембран имеются специфические структурные образования, названные Парком квантосомами.
Хлоропласты – системы, способные к автономному синтезу белков. В них присутствуют низко- и высокомолекулярная РНК, специфическая кольцевая ДНК и ферменты, активирующие аминокислоты. Хлоропласты обладают собственными рибосомами.
1) осуществление фотосинтеза (основная функция);
2) в хлоропластах происходит вся сложная цепь процессов превращений первичных продуктов фотосинтеза (наращивание углеродной цепи, образование и распад полимерных форм углеводов и др.);
Биохимические системы синтеза и превращения углеводов функционируют в строме хлоропластов. В ней же может откладываться крахмал. В хлоропластах представлен весь набор биохимических систем, участвующих в синтезе АТФ.
Онтогенез хлоропластов. При нормальном освещении пропластиды пре-вращаются в хлоропласты. Сначала они растут, при этом происходит образование продольно расположенных мембранных складок от внутренней мембраны. Одни из них простираются по всей длине пластиды и формируют ламеллы стромы; другие образуют ламеллы тилакоидов, которые выстраиваются в виде стопки и образуют граны зрелых хлоропластов.
Несколько иначе развитие пластид происходит в темноте. У этиолированных проростков происходит в начале увеличение объема пластид – этиопластов, но система внутренних мембран не строит ламеллярные структуры, а образует массу мелких пузырьков, которые скапливаютсяя в отдельные зоны и даже могут формировать сложные решетчатые структуры (проламеллярные тела). В мембранах этиопластов содержится протохлорофилл, предшественник хлорофилла желтого цвета. Под действие света из этиопластов образуются хлоропласты, протохлорофилл превращается в хлорофилл, происходит синтез новых мембран, фотосинтетических ферментов и компонентов цепи переноса электронов.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом