
Прежде всего глюкоза подвергается фосфорилированию при участии фермента гексокиназы, а в печени – и глюкокиназы. Далее глюкозо-6-фосфат под влиянием фермента фосфоглюкомутазы переходит в глюкозо-1-фос-фат:
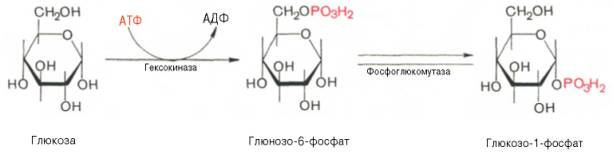
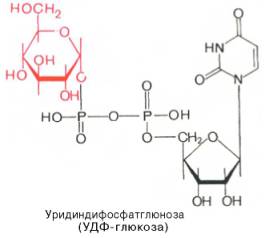
Образовавшийся глюкозо-1-фосфат уже непосредственно вовлекается в синтез гликогена. На первой стадии синтеза глюкозо-1-фосфат вступает во взаимодействие с УТФ (уридинтрифосфат), образуя уридиндифосфатглюкозу (УДФ-глюкоза) и пирофосфат. Данная реакция катализируется ферментом глюкозо-1-фосфат-уридилилтрансферазой (УДФГ-пирофосфорилаза):
Глюкозо-1-фосфат + УТФ УДФ-глюкоза + Пирофосфат.
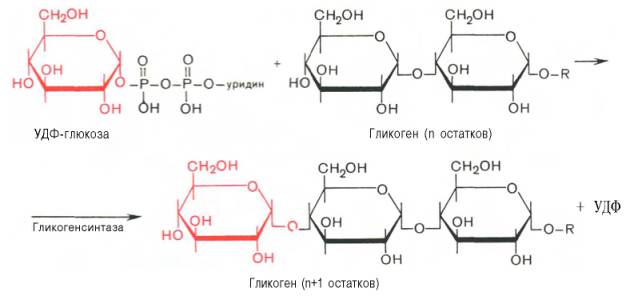
На второй стадии – стадии образования гликогена – происходит перенос глюкозного остатка, входящего в состав УДФ-глюкозы, на глюкозидную цепь гликогена («затравочное» количество). При этом образуется α-(1–>4)-связь между первым атомом углерода добавляемого остатка глюкозы и 4-гидроксильной группой остатка глюкозы цепи. Эта реакция катализируется ферментом гликогенсинтазой. Необходимо еще раз подчеркнуть, что реакция, катализируемая гликогенсинтазой, возможна только при условии, что полисахаридная цепь уже содержит более 4 остатков D-глю-козы.
Образующийся УДФ затем вновь фосфорилируется в УТФ за счет АТФ, и таким образом весь цикл превращений глюкозо-1-фосфата начинается сначала.
В целом образование α-1,4-глюкозидной ветви («амилозной» ветви) гликогена можно представить в виде следующей схемы:
Установлено, что гликогенсинтаза неспособна катализировать образование α-(1–>6)-связи, имеющейся в точках ветвления гликогена. Этот процесс катализирует специальный фермент, получивший название гли-когенветвящего фермента, или амило-(1–>4)–>(1–>6)-трансглюкозидазы. Последний катализирует перенос концевого олигосахаридного фрагмента, состоящего из 6 или 7 остатков глюкозы, с нередуцирующего конца одной из боковых цепей, насчитывающей не менее 11 остатков, на 6-гидроксиль-ную группу остатка глюкозы той же или другой цепи гликогена. В результате образуется новая боковая цепь.
Ветвление повышает растворимость гликогена. Кроме того, благодаря ветвлению создается большое количество невосстанавливающих концевых остатков, которые являются местами действия гликогенфосфорилазы и гликогенсинтазы.
Таким образом, ветвление увеличивает скорость синтеза и расщепления гликогена.
Благодаря способности к отложению гликогена (главным образом в печени и мышцах и в меньшей степени в других органах и тканях) создаются условия для накопления в норме некоторого резерва углеводов. При повышении энерготрат в организме в результате возбуждения ЦНС обычно происходят усиление распада гликогена и образование глюкозы.
Помимо непосредственной передачи нервных импульсов к эффекторным органам и тканям, при возбуждении ЦНС повышаются функции ряда желез внутренней секреции (мозговое вещество надпочечников, щитовидная железа, гипофиз и др.), гормоны которых активируют распад гликогена, прежде всего в печени и мышцах (см. главу 8).
Как отмечалось, эффект катехоламинов в значительной мере опосредован действием цАМФ, который активирует протеинкиназы тканей. При участии последних происходит фосфорилирование ряда белков, в том числе гликогенсинтазы и фосфорилазы b – ферментов, участвующих в обмене углеводов. Фосфорилированный фермент гликогенсинтаза сам по себе малоактивен или полностью неактивен, но в значительной мере активируется положительным модулятором глюкозо-6-фосфатом, который увеличивает Vmaxфермента. Эта форма гликогенсинтазы называется D-формой, или зависимой (dependent) формой, поскольку ее активность зависит от глюкозо-6-фосфата. Дефосфорилированная форма гликоген-синтазы, называемая также I-формой, или независимой (independent) формой, активна и в отсутствие глюкозо-6-фосфата.
Таким образом, адреналин оказывает двойное действие на обмен углеводов: ингибирует синтез гликогена из УДФ-глюкозы, поскольку для проявления максимальной активности D-формы гликогенсинтазы нужны очень высокие концентрации глюкозо-6-фосфата, и ускоряет распад гликогена, так как способствует образованию активной фосфорилазы а. В целом суммарный результат действия адреналина состоит в ускорении превращения гликогена в глюкозу.
Биосинтез гликогена
Расщепление и синтез гликогена осуществляют различные ферменты.
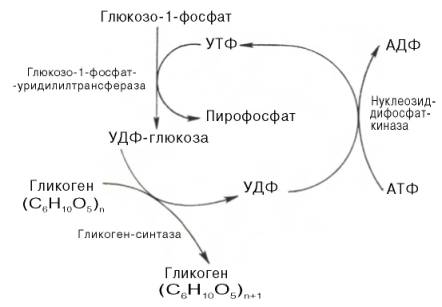
Расщепление гликогена с образованием глюкозо-1-фосфата катализирует гликоген-фосфорилаза (рис. 5). А синтез гликогена происходит при участии других ферментов.
Основные этапы синтеза гликогена следующие.
1. На первом этапе из свободной глюкозы образуется глюкозо-6-фосфат:
АТФ + Глюкоза > Глюкозо-6-фосфат + АДФ
2. Затем из глюкозо-6-фосфата образуется глюкозо-1-фосфат. Эта реакция обратима.
3. Следующий этап является ключевым. В реакции катализируемой глюкозо-1-фосфат-уридилтрансферазой образуется уридилдифосфатглюкоза (UDP-глюкоза):
UТP + Глюкозо-1-фосфат > UDP-глюкоза + РРi.
4. Далее гликоген-синтаза катализирует перенос гликозильных групп от UDP-глюкозы на молекулу гликогена с образованием альфа(1-4)-связи.
5. Образование боковой цепи гликогена катализирует «ветвящий» фермент гликозил-(4-6)-трансфераза.
В организме человека, как и животных, гликоген синтезируется практически во всех тканях, но больше всего в печени и скелетных мышцах.
Регуляция расщепления и синтеза гликогена также взаимосвязана
Также как гликолиз и глюконеогенез, расщепление и синтез гликогена регулируются взаимосвязано.
Если гликоген-фосфорилаза активируется, то гликоген-синтаза ингибируется, и наоборот.
Регуляцию этих ферментов в печени в конечном счете осуществляют гормоны: инсулин и контринсулярные гормоны, например, адреналин и глюкагон. Инсулин ингибирует распад и стимулирует синтез гликогена.
Читайте также: Ткани идеальные для лета
Контринсулярные гормоны стимулируют расщепление гликогена. Некоторые контринсулярные гормоны проявляют селективность по отношению к различным органам и тканям. Например, глюкагон стимулирует расщепление гликогена только в печени, но не в скелетных мышцах.
Гликогенозы. Ряд наследственных болезней связан с нарушением обмена гликогена. Эти болезни получили название гликогенозов. Они возникают в связи с дефицитом или полным отсутствием ферментов, катализирующих процессы распада или синтеза гликогена, и характеризуются избыточным его накоплением в различных органах и тканях (табл. 10.2).
Гликогеноз I типа (болезнь Гирке) встречается наиболее часто, обусловлен наследственным дефектом синтеза фермента глюкозо-6-фосфатазы впечени и почках. Болезнь наследуется по аутосомно-рецессивному типу. Патологические симптомы появляются уже на первом году жизни ребенка: увеличена печень, нередко увеличены почки. В результате гипогликемии появляются судороги, задержка роста, возможен ацидоз. В крови – повышенное количество лактата и пирувата. Введение адреналина или глюка-гона вызывает значительную гиперлактатацидемию, но негипергликемию, так как глюкозо-6-фосфатаза в печени отсутствует и образования свободной глюкозы не происходит.
57. Регуляция обмена углеводов и поддержания физиологического уровня глюкозы в крови. Характеристика гормонов и механизм их действия. Гипо- и гипергликемии.
. Гормоны. К гормонам, которые влияют на углеводный обмен, принадлежат пептиды инсулин иглюкагон, глюкокортикоид кортизол и катехоламин адреналин (см. сс. 362, 368). Инсулининдуцирует (см. с. 120) синтез de novo гликоген-синтазы [1], а также некоторых ферментов гликолиза [3, 5, 7]. Одновременно инсулин подавляет синтез ключевых ферментов глюконеогенеза (репрессия,[4, 6, 8, 9]). Глюкагон как антагонист инсулина действует в противоположном направлении: индуцирует ферменты глюконеогенеза [4, 6, 8, 9] и репрессирует пируваткиназу [7], ключевой фермент гликолиза. Другие аффекты глюкагона основаны на взаимопревращении ферментов и опосредованы вторичным мессенджером цАМФ (сАМР, см. с. 114). По этому механизму тормозится синтез гликогена [1] и активируется расщепление гликогена [2]. Подобным образом действует и адреналин. Торможение пируваткиназы [7] глюкагоном также обусловлено взаимопревращением ферментов.
Глюкокортикоиды, прежде всего кортизол (см. с. 362), индуцируют все ключевые ферменты глюконеогенеза [4, 6, 8, 9]. Одновременно они индуцируют ферменты деградации аминокислот и обеспечивают тем самым глюконеогенез исходными соединениями.
Метаболиты. Высокие концентрации АТФ (АТР) и цитрата тормозят гликолиз путем аллостерической регуляции фосфофруктокиназы. Кроме того, АТФ тормозит пируваткиназу. Ингибитором пируваткиназы является ацетил-КоА. Все эти метаболиты образуются при распаде глюкозы (торможение конечным продуктом). АМФ (AMP), сигнал дефицита АТФ, активирует расщепление гликогена и тормозит глюконеогенез.
Гипогликемия. Нередко гипогликемия связана с понижением функций тех эндокринных желез, повышение функций которых приводит, как отмечалось, к гипергликемии. В частности, гипогликемию можно наблюдать при гипофизарной кахексии, аддисоновой болезни, гипотиреозе. Резкое снижение уровня глюкозы в крови отмечается при аденомах поджелудочной железы вследствие повышенной продукции инсулина β-клетками панкреатических островков. Кроме того, гипогликемия может быть вызвана голоданием, продолжительной физической работой, приемом β-ганглиоблока-торов. Низкий уровень глюкозы в крови иногда отмечается при беременности, лактации.
Гипогликемия может возникнуть при введении больным сахарным диабетом больших доз инсулина. Как правило, она сопровождает почечную глюкозурию, возникающую вследствие снижения «почечного порога» для глюкозы.
58. Особенности обмена глюкозы в различных тканях (мышцы, эритроциты, мозг, жировая ткань, печень). Зависимость путей использования глюкоза от ритма и характера питания.
ОСОБЕННОСТИ И ЗНАЧЕНИЕ ГЛИКОЛИЗА В ЭРИТРОЦИТАХ 1. Генерация АТФ. АТФ используется для 1) активного транспорта катионов через мембрану, 2) сохранения целостности мембраны и формы эритроцитов. 2. В процессе гликолиза генерируется НАДН2, который является: 1) кофактором метгемоглобинредуктазы — фермента, катализирующего переход мет-Hb в Hb; этот процесс предотвращает накопление мет-Hb (кофактором может являться и НАДФН2); 2) кофактором ЛДГ (лактатдегидрогеназы); 3) поставщиком протонов для супероксиддисмутазной реакции. 3. В процессе гликолиза 1,3-дифосфоглицерат (1,3-ДФГ) превращается в 2,3-ДФГ. На этот процесс расходуется 20-25% глюкозы. 2,3-ДФГ — активная отрицательно заряженная молекула. В эритроцитах периферической крови образует солевую связь с Hb, уменьшает его сродство к кислороду, что обеспечивает переход кислорода в клетки тканей. В капиллярах легких Hb освобождается от 2,3-ДФГ и приобретает способность акцептировать кислород. ПФП — ГЛАВНОЕ НАЗНАЧЕНИЕ — генерация восстановленного кофактора НАДФН2, который используется в эритроцитах для восстановления глутатиона при участии глутатионредуктазы, поставляет протоны для супероксидодисмутазной реакции, хотя и слабо, но используется мет-Hb-редуктазой для восстановления мет-Hb в Hb. Промежуточный продукт ПФП — 3-ФГА (3-фосфоглицериновый альдегид) используется в процессе гликолиза, в том числе и для синтеза 2,3-ДФГ.
Читайте также: Основная масса языка ткань
Глюкоза высвобождается из печени в промежутках между приемами пищи. Если уровень глюкозы в крови начинает снижаться до нижнего предела в промежутках между приемами пищи, это приведен к ряду изменений и послужит поводом для высвобождения глюкозы из печени в кровоток. 1. Снижение уровня глюкозы приводит к снижению секреции инсулина поджелудочной железой.
2. Отсутствие инсулина приведет к изменению направления реакций, нацеленных на создание запаса гликогена, главным образом к остановке дальнейшего синтеза гликогена в печени и предупреждению поступления глюкозы в печень из крови.
3. Отсутствие инсулина (параллельно с увеличением глюкагона, что будет рассмотрено далее) активирует фермент фосфорилазу, расщепляющую гликоген до глюкозофосфата.
4. Фермент глюкофосфатаза, ингибируемый инсулином, при отсутствии инсулина активируется и приводит к отщеплению фосфатного радикала от глюкозы, что позволяет свободной глюкозе вернуться в кровь.
Таким образом, печень забирает глюкозу из крови, когда в крови возникает ее избыток в связи с приемом пищи, и возвращает ее в кровь, когда концентрация глюкозы снижается в промежутках между приемами пищи. Обычно около 60% глюкозы пищи запасается таким способом в печени и в последующем возвращается в кровь. Инсулин обеспечивает превращение избытка глюкозы в жирные кислоты и ингибирует глюконеогенез в печени.
Цель мышечной клетки – наиболее эффективно использовать поступающую глюкозу для образования АТФ, необходимого для осуществления механической работы – сокращения. В состоянии покоя значительные количества глюкозы резервируются в форме гликогена. Цитоплазма мышечных клеток содержит в высоких концентрациях ферменты гликолиза, а изобилие митохондрий обеспечивает эффективный распад продуктов гликолиза через путь лимонной кислоты и цепь переноса электронов. Лишь в условиях крайнего утомления эти аэробные процессы е справляются с накоплением лактата.
В мышцах идет гликогенез, мышца осуществляет лишь немногие синтетические функции. Ключевые ферменты глюконеогенеза в мышцах отсутствуют, и глюконеогенез не идет. Для востановительных синтезов в мышце НАДФ . Н не требуется, и фосфоглюконатный путь почти не функционирует.
Обмен углеводов в мышцах обеспечивает создание тканевых запасов гликогена в состоянии покоя и использование этих запасов, а также поступающей глюкозы при напряженной работе; основные энергетические потребности всех типов мышц удовлетворяются главным образом за счет окисления продуктов обмена жиров. Ни медленно сокращающаяся гладкая мышечная ткань, ни сердечная мышца не потребляют глюкозу в значительной мере. Во время напряженной работы сердце обеспечивает себя лактатом для окисления.
Синтез гликогена (гликогеногенез)
Синтез гликогена (гликогеногенез)
Гликоген синтезируется в период пищеварения (через 1–2 часа после приема углеводный пищи). Синтез гликогена из глюкозы, как и любой анаболический процесс, является эндергоническим, т. е. требует затрат энергии.
Синтез гликогена включает 4 этапа:
1. Фосфорилирование глюкозы до глюкозо-6-фосфата при участии гексокиназы или глюкокиназы.
2. Активация первого углеродного атома с образованием активной формы – УДФ – глюкозы.
3. Образование ?-1,4-гликозидных связей. В присутствии «затравки» гликогена (молекулы, включающей не менее 4 остатков глюкозы) фермент гликогенсинтаза присоединяет остатки глюкозы из УДФ-глюкозы к С4-атому концевого остатка глюкозы в гликогене, образуя ?-1,4-гликозидную связь.
4. Образование ?-1,6-гликозидных связей (точки ветвления молекулы). Образование их осуществляется амилозо-1,4 ? 1,6-трансглюкозидазой (ветвящий или бранчинг фермент). Когда длина линейного участка цепи включает минимально 11 остатков глюкозы, этот фермент переносит фрагмент (1 ? 4) цепи с минимальным количеством 6 остатков глюкозы на соседнюю цепь или на несколько участков глюкозы дальше, образуя ?-1,6-гликозидную связь. Таким образом, образуется точка ветвления. Ветви растут путем последовательного присоединения (1–4)-глюкозильных единиц и дальнейшего ветвления.
Гликогенсинтаза – регуляторный фермент, существующий в двух формах:
1. – дефосфорилированной, активной (форма а);
2. – фосфорилированной, неактивной (форма b).
Активная форма образуется из неактивной под действием фосфатазы гликогенсинтазы при дефосфорилировании. Превращение активной формы в неактивную происходит при участии протеинкиназы путем фосфорилирования за счет АТФ.

Рис. 18.-1. Регуляция активности гликогенсинтазы.
Распад гликогена может проходить двумя путями.
Читайте также: Обеспечивает твердость частям растения какая ткань
1. Гидролитический – при участии амилазы с образованием декстринов и даже свободной глюкозы.
2. Фосфоролитический – под действием фосфорилазы и образованием глюкозо-1-фосфата. Это основной путь распада гликогена.
Фосфорилаза – сложный регуляторный фермент, существующий в двух формах – активной и неактивной. Активная форма (фосфорилаза а) – это тетрамер, в котором каждая субъединица соединена с остатком ортофосфата через гидроксильную группу серина. Под действием фосфатазы фосфорилазы происходит дефосфорилирование, отщепление 4 молекул фосфорной кислоты, и фосфорилаза а превращается в неактивную форму – фосфорилазу b, распадаясь на две димерные молекулы. Фосфорилаза b активируется путем фосфорилирования остатков серина за счет АТФ под действием фермента киназы фосфорилазы. В свою очередь этот фермент также существует в двух формах. Активная киназа фосфорилазы – фосфорилированный фермент, превращается в неактивную форму под действием фосфатазы. Активация киназы фосфорилазы осуществляется путем фосфорилирования за счет АТФ в присутствии ионов Mg 2+ протеинкиназой.
Регуляция синтеза и распада гликогена носит каскадный характер и происходит путем химической модификации ферментов.
Поскольку синтез и распад гликогена протекают по разным метаболическим путям, эти процессы могут контролироваться реципрокно. Влияние гормонов на синтез и распад гликогена осуществляется путем изменения в противоположных направлениях активности двух ключевых ферментов: гликогенсинтазы и гликогенфосфорилазы с помощью их фосфорилирования и дефосфорилирования. Инсулин стимулирует синтез гликогена и тормозит распад, адреналин и глюкагон обладают противоположным эффектом.
Данный текст является ознакомительным фрагментом.
Читайте также
5. Синтез рибосомных РНК
5. Синтез рибосомных РНК В обычных клетках синтез трех видов рРНК (28S, 18S и малой 5S) координирован, т. е. на одну молекулу 28S образуется одна молекула 18S и одна молекула 5S. Синтез 28S и 18S происходит в виде одного большого, общего для них предшественника (пре-рРНК), который затем
6. Гормоны регулируют синтез желтка и белка
6. Гормоны регулируют синтез желтка и белка Мы уже говорили, что у позвоночных животных желток будущего яйца синтезируется в печени. Этот синтез стимулируется женскими половыми стероидными гормонами — эстрогенами (подробнее см. специальную главу). Один из таких гормонов
Великий синтез
Великий синтез Как связать эволюцию с генетикой. Можно ли подойти к вопросам изменчивости, борьбы за существование, отбора — словом, дарвинизма, исходя не из тех совершенно бесформенных, расплывчатых, неопределенных воззрений на наследственность, которые только и
2.4. Конфронтация или новый синтез?
2.4. Конфронтация или новый синтез? Наиболее оправданной позицией многим эволюционистам уже давно представлялся синтез положений СТЭ с концепциями направленной эволюции и сальтационизма на основе достижений генетики. Разные авторы говорили, что пора переходить от
3. ПОВТОРНЫЙ ВХОД ВОЗБУЖДЕНИЯ И ИНФОРМАЦИОННЫЙ СИНТЕЗ
3. ПОВТОРНЫЙ ВХОД ВОЗБУЖДЕНИЯ И ИНФОРМАЦИОННЫЙ СИНТЕЗ Описанная ранее концепция «светлого пятна» исходит из того, что сознание определяется некоторым уровнем возбудимости мозговых структур. Однако можно предположить, что этого недостаточно и в действительности
Глава 18. Обмен гликогена
Глава 18. Обмен гликогена Гликоген – основной резервный полисахарид в животных тканях. Он представляет собой разветвленный гомополимер глюкозы, в котором остатки глюкозы соединены в линейных участках ?-1,4-гликозидными связями, а в точках ветвления – ?-1,6- гликозидными
Нарушения обмена гликогена
Нарушения обмена гликогена Гликогеновые болезни – группа наследственных нарушений в основе которых лежит снижение или отсутствие активности ферментов, катализирующих реакции синтеза или распада гликогена. К данным нарушениям относятся гликогенозы и
Синтез жирных кислот
Синтез жирных кислот Синтез жирных кислот происходит в основном в печени, в меньшей степени – в жировой ткани и лактирующей молочной железе. Гликолиз и последующее окислительное декарбоксилирование пирувата способствуют увеличению концентрации ацетил-КоА в матриксе
5.5. Альтернативные теории и синтез идей эволюционизма
5.5. Альтернативные теории и синтез идей эволюционизма В рамках научной методологии эволюционизму нет альтернативы, поскольку такой альтернативой может служить только креационизм. Однако сам эволюционизм не является однородным течением. Хотя после чтения популярных
Передача гормонального сигнала: синтез, секреция, транспорт гормонов, их действие на клетки-мишени и инактивация
Передача гормонального сигнала: синтез, секреция, транспорт гормонов, их действие на клетки-мишени и инактивация В определении понятия «гормон» было указано несколько этапов распространения гормонального сигнала (рис. 2.6). Рис. 2.6. Этапы распространения гормонального
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом
- Правообладателям
- Политика конфиденциальности