
Основные органы ткани мишени для гормона инсулина это
а) Активация инсулином рецепторов клеток-мишеней и результирующие ответы клеток. Для воздействия на клетку-мишень инсулин прежде всего связывается и активирует рецептор, расположенный на мембране клеток, — белок с молекулярной массой около 300000 (для облегчения понимания просим вас изучить рисунок ниже).
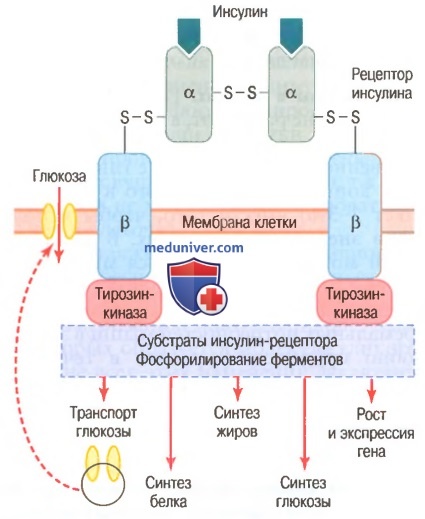
Схема инсулин-рецептора. Инсулин связывается с альфа-субъединицей рецептора, что вызывает фосфорилирование бета-субъединицы рецептора.
Это, в свою очередь, индуцирует активность тирозинкиназы. Рецептор тирозинкиназной активности запускает каскад реакций фосфорилирования в клетке, что увеличивает или снижает активность ферментов, включая субстраты инсулин-рецептора, которыми опосредованы влияния глюкозы на метаболизм глюкозы, жиров и белков.
Например, транспортеры глюкозы продвигаются к мембране клетки, облегчая поступление глюкозы в клетку
Активация рецептора сопровождается определенной последовательностью реакций.
Рецептор инсулина состоит из 4 субъединиц, связанных дисульфидными связями: 2 альфа-субъединицы располагаются практически снаружи мембраны и 2 бета-субъединицы прободают мембрану и продолжаются в цитоплазму. Инсулин взаимодействует с 2 наружными субъединицами, но благодаря существенным собственным связям субъединиц между собой внутренние бета-субъединицы при этом аутофосфорилируются, поэтому инсулиновый рецептор является примером энзим-связанного рецептора.
Аутофосфорилирование бета-субъединиц рецептора приводит к местной активации тирозинкиназы, которая в итоге вызывает фосфорилирование многих других внутриклеточных ферментов, включая группу, названную субстратами инсулин-рецептора. Различные типы СИР (например, СИР1, СИР2, СИРЗ) представлены в различных клетках. Система действует, активируя одни ферменты одновременно с инактивацией других. Таким образом, инсулин управляет внутриклеточными метаболическими механизмами, вызывая желательные реакции обмена белков, жиров и углеводов. Окончательные эффекты действия инсулина следующие.
1. Через несколько секунд после взаимодействия инсулина с рецептором проницаемость мембран для глюкозы у 80% клеток организма заметно повышается. Это особенно справедливо для мышечных клеток и адипоцитов, но совершенно не распространяется на нейроны мозга. Поступившая в клетки глюкоза фосфорилируется и становится субстратом для всех обычных этапов обмена углеводов.
Увеличение транспорта глюкозы в клетку предположительно является результатом перемещения множества внутриклеточных пузырьков к мембране клетки; эти пузырьки несут на своей мембране множество белков, обеспечивающих транспорт глюкозы; взаимодействуя с мембраной клетки, они облегчают поступление глюкозы в клетку. Если инсулина уже не оказывается в наличии, пузырьки отделяются от мембраны в течение 3-5 мин и возвращаются в середину клетки, сохраняя способность к использованию их по мере необходимости.
2. Мембрана становится более проницаемой для многих аминокислот, ионов калия, фосфатов, повышая транспорт этих веществ в клетку.
3. Медленные эффекты возникают через 10-15 мин и заключаются в изменении уровня активности большинства внутриклеточных метаболических ферментов. Этот эффект является результатом главным образом изменения процессов фосфорилирования ферментов.
4. Более медленные процессы продолжаются в течение нескольких часов и дней. Они являются результатом изменения скорости процесса трансляции на рибосомах и приводят к образованию новых белков. Еще более медленные эффекты опосредованы изменением скорости транскрипции ДНК в ядре клетки. Таким способом инсулин формирует многие ферментативные механизмы клеток для выполнения свойственных им метаболических задач.
Видео физиология гормонов поджелудочной железы и регуляции уровня глюкозы — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Читайте также: Почему кожицу лука можно назвать тканью
Основные органы ткани мишени для гормона инсулина это
Действие инсулина на клетки-мишени начинается после его связывания со специфическими димерными мембранными рецепторами (рис. 6.22), при этом внутриклеточный домен рецептора обладает тирозинкиназной активностью. Инсулин-рецепторный комплекс не только передает сигнал внутрь клетки, но и частично путем эндоцитоза поступит внутрь клетки к лизо-сомам. Под влиянием лизосомальной протеазы инсулин отщепляется от рецептора, при этом последний либо разрушается, либо возвращается к мембране и вновь встраивается в нее. Многократное перемещение рецептора от мембраны к лизосомам и обратно к мембране носит название рециклизация рецептора. Процесс рециклизации важен для регуляции колич-ства инсулиновых рецепторов, в частности обеспечения обратной зависимости между концентрацией инсулина и количеством мембранных рецепторов к нему.
Образование инсулин-рецепторного комплекса активирует тирозинкиназу, запускающую процессы фосфорилирования внутриклеточных белков. Происходящее при этом аутофосфорилирование рецептора ведет к усилению первичного сигнала. Инсулин-рецепторный комплекс вызывает активирование фосфолипазы С, образование вторичных посредников инозитолтрифосфата и диацилглицерола, активацию протеинкиназы С, ингибирование цАМФ. Участие нескольких систем вторичных посредников объясняет многообразие и различия эффектов инсулина в разных тканях.
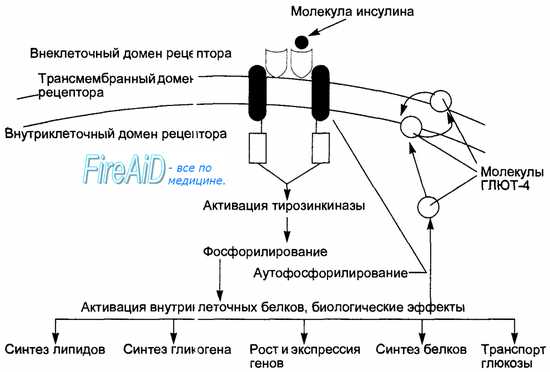
Рис. 6.22. Схема механизма действия инсулина на клетку-мишень.
Инсулин оказывает влияние на все виды обмена веществ, способствует анаболическим процессам, увеличивая синтез гликогена, жиров и белков, тормозя эффекты многочисленных контринсулярных гормонов (глюкагона, катехоламинов, глюкокортикоидов и соматотропина). Все эффекты инсулина по скорости их реализации подразделяют на 4 группы: очень быстрые (через несколько секунд) — гиперполяризация мембран клеток (за исключением гепатоцитов), повышение проницаемости для глюкозы, активация Na-K-АТФазы, входа К+ и откачивания Na , подавление Са-насоса и задержка Са2+; быстрые эффекты (в течение нес кольких минут) — активация и торможение различных ферментов, подавляющих катаболизм и усиливающих анаболические процессы; медленные процессы (в течение нескольких часов) — повышенное поглощение амиминокислот, изменение синтеза РНК и белков-ферментов; очень медленные эффекты (от часов до суток) — активация митогенеза и размножения клеток.
Важнейшим эффектом инсулина в организме является увеличение в 20—50 раз транспорта глюкозы через мембраны мышечных и жировых клеток путем облегченной диффузии по градиенту концентрации с помощью чувствительных к гормон) мембранных белковых переносчиков, называемых ГЛЮТ. В мембранах разных видов клеток выявлены 6 типов ГЛЮТ (рис. 6.23), но только один из них — ГЛЮТ-4 — является инсулинозависимым и находится в мембранах клеток скелетных мышц, миокарда, жировой ткани.
Инсулин влияет на угле водный обмен, что проявляется:
1) активацией утилизации глюкозы клетками,
2) усилением процессов фосфорилирования;
3) подавлением распад; и стимуляцией синтеза гликогена;
4) угнетением глюконеогенеза;
5) активацией процессов гликолиза;
6) гипогликемией.
Действие инсулина на белковый обмен состоит в: 1) повышении проницаемости мембран для аминоокислот; 2) усилении синтеза иРНК; 3) активации в печени синтеза aминокислот; 4) повышении синтеза и подавлении распада белка.
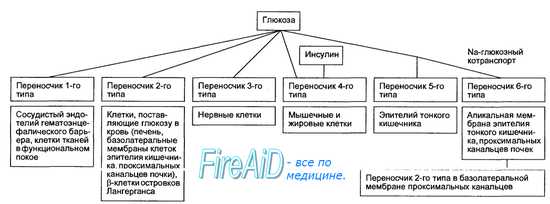
Рис. 6.23. Схема транспорта глюкозы через клеточные мембраны. Переносчики имеют общее название ГЛЮТ-1, 2, 3, 4, 5, 6. Только ГЛЮТ-4 является инсулинозависимым.
Читайте также: Бархат ткань какое оно
Основные эффекты инсулина на липидный обмен:
• стимуляция синтеза свободных жирных кислот из глюкозы;
• стимуляция синтеза липопротеиновой липазы в клетках эндотелия сосудов и благодаря этому активация гидролиза связанных с липо-протеинами крови триглицеридов и поступления жирных кислот в клетки жировой ткани;
• стимуляция синтеза триглицеридов;
• подавление распада жира;
• активация окисления кетоновых тел в печени.
Благодаря влиянию на клеточную мембрану инсулин поддерживает высокую внутриклеточную концентрацию ионов калия, что необходимо для обеспечения нормальной в возбудимости клеток.
Широкий спектр метаболических эффектов инсулина в организме свидетельствует о том, что гормон необходим для осуществления функционирования всех тканей, оргаганов и физиологических систем, реализации эмоциональных и поведенческих актов, поддержания гомеостазиса, осуществления механизмов приспособления и защиты организма от неблагоприятных факторов среды.
Недостаток инсулина (относительный дефицит по сравнению с уровнем контринсулярных гормонов, прежде всего глюкагона) приводит к сахарному диабету. Избыток инсулина в крови, например при передозировке, вызывает гипогликемию с резкими нарушениями функций центральной нервной системы, использующей глюкозу как основной источник энергии независимо от инсулина.
Поговорим про инсулин
Инсулин — это гормон поджелудочной железы, который главным образом воздействует на обмен веществ, причем в основном — на концентрацию глюкозы в крови. В своих тканях-мишенях он влияет как на мембранные, так и на внутриклеточные процессы. Некоторые из его эффектов перечислены в ниже.
Эффекты инсулина

Мембранные эффекты
- Стимуляция транспорта глюкозы (и некоторых других моносахаридов)
- Стимуляция транспорта аминокислот (особенно аргинина)
- Стимуляция транспорта жирных кислот
- Стимуляция поглощения клеткой К+ и Mg2+
Внутриклеточные эффекты
- Стимуляции синтеза РНК и ДНК
- Стимуляция синтеза белка
- Усиленная стимуляция гликогенсинтазы (гликогенез)
- Стимуляция глюкокиназы
- Ингибирование глюкозо-6-фосфатазы
- Стимуляция липогенеза
- Ингибирование липолиза (ингибирование синтеза цАМФ)
- Стимуляция синтеза жирных кислот
- Активация Mg2+-стимулируемой Na+/K+-АТФазы
Механизм действия инсулина и влияние его на обмен

Инсулин и глюкоза
Попав в клетку, глюкоза быстро превращается в глюкозо-6-фосфат, поэтому ее внутриклеточная концентрация остается крайне низкой. Уровень глюкозы в артериальной крови в норме поддерживается в пределах 4-8 ммоль/л (72-144 мг/100 мл), так что по обе стороны клеточной мембраны всегда существует градиент ее концентраций. Несмотря на это, однако, простая диффузия обеспечивает поступление в большинство клеток лишь небольшого количества глюкозы, которого явно недостаточно для удовлетворения их метаболических потребностей (даже при возрастании концентрационного градиента, как это имеет место при высокой гипергликемии). В присутствии же инсулина проникновение декстрозы в клетки резко усиливается. Это действие инсулина проявляется лишь при наличии концентрационного градиента глюкозы, конкурентно ингибируется другими моносахаридами (например, галактозой) и следует кинетике насыщаемого процесса. Таким образом, гормон стимулирует процесс облегченной диффузии декстрозы, который осуществляется при участии чувствительных к гормону белковых транспортеров глюкозы (GLUT), расположенных на клеточной мембране. Эти транспортеры способны переносить глюкозу через клеточную мембрану в обоих направлениях, но ее поток зависит от концентрационного градиента, который направлен из внеклеточного пространства во внутриклеточное. В разных клетках найдены многочисленные GLUT, но инсулинозависимым является только один из этих белков — GLUT4, и именно он присутствует в мембранах клеток скелетных и сердечных мышц, а также жировой ткани.
Читайте также: Артефакт ткани venus premium
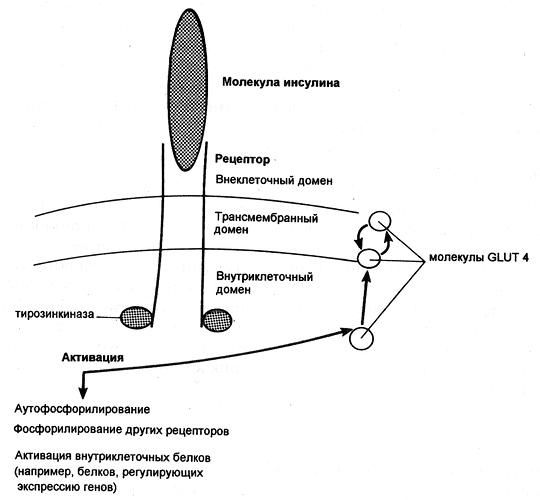
Димерный рецептор инсулина и последствия инсулиновой активации тирозинкиназы (GLUT — транспортер глюкозы)
Некоторые ткани полностью удовлетворяют свои потребности в глюкозе за счет инсулиннезависимых механизмов. Например, в клетки печени и центральной нервной системы декстроза попадает с помощью инсулиннезависимых GLUT, и поглощение этими тканями зависит только от ее уровня в крови. Кроме того, мембрану эритроцитов, клеток почек и кишечника глюкоза пересекает вместе с ионами натрия, которые поступают в клетки путем пассивной диффузии по градиенту концентрации.
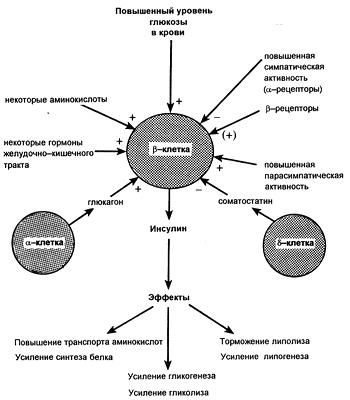
Регуляция продукции инсулина
Инсулин влияет и на внутриклеточные процессы обмена веществ. В печеночных и других клетках он стимулирует синтез гликогена, повышая активность гликогенсинтазы, что ускоряет включение гликозильных остатков в гликоген. Гормон поджелудочной железы повышает также активность печеночной глюкокиназы; этот фермент катализирует фосфорилирование глюкозы (с образованием глюкозо-6-фосфата). Одновременно гормон ингибирует печеночную фосфатазу, которая дефосфорилирует глюкозо-6-фосфат, с образованием свободной глюкозы. Такие изменения активности печеночных ферментов обусловливают снижение продукции декстрозы и наряду со стимуляцией поглощения ее периферическими клетками определяют гипогликемию, возникающее под влиянием инсулина. Возрастающая под действием последнего утилизация глюкозы в тканях обеспечивает сохранение запасов других внутриклеточных энергетических субстратов, таких как жиры и белки.
Белки и инсулин
Инсулин стимулирует не только активный транспорт аминокислот в периферические клетки, но и непосредственно синтез белка. Поскольку эти два эффекта могут не зависеть друг от друга, гормон влияет, очевидно, не только на клеточную мембрану, но и на внутриклеточные процессы. Стимуляции синтеза белка предшествует возрастание активности мРНК. Поскольку гормон с трудом проходит сквозь мембраны клеток, в механизме его ядерного эффекта должен принимать участие второй посредник. Синтез белка под действием инсулина усиливается и вследствие возрастания количества поступающих в клетку аминокислот. С другой стороны, возрастание утилизации глюкозы замедляет распад белка. Ускорение синтеза и замедление распада белка под влиянием гормона приводят к увеличению белковых запасов в интрацеллюлярном секторе.
Все эти эффекты определяют важнейшую роль инсулина в регуляции процессов роста и развития.
Инсулин и жир
Инсулин стимулирует поглощение и окисление глюкозы клетками жировой ткани. Он также стимулирует синтез липопротеиновой липазы в эндотелиальных клетках. Этот фермент катализирует гидролиз триглицеридов, связанных с липопротеинами крови, и способствует поступлению жирных кислот в адипоциты. Наряду с прямой стимуляцией липогенеза в печени и жировой ткани это приводит к увеличению запасов жира. Кроме того, инсулин ингибирует опосредуемый цАМФ липолиз, тормозя гормончувствительную внутриклеточную липопротеиновую липазу.
Инсулин и калий
Присутствие инсулина необходимо для поддержания внутриклеточной концентрации ионов калия; этот эффект, по всей вероятности, является следствием прямого влияния гормона на клеточную мембрану.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом