
Всякая живая клетка обладает свойствами раздражимости, возбудимости и лабильности (функциональной подвижности).
Раздражимость — общее свойство любой живой ткани или клетки реагировать на раздражение изменением обмена веществ и энергии. Этим свойством (раздражимостью) обладают как животные, так и растения, низшие и высшие их формы. Раздражимость лежит в основе постепенного морфологического и функционального приспособления отдельных тканей или всего организма к длительным изменениям внешней и внутренней среды. Раздражимость присуща всем тканям, в том числе и таким высокоорганизованным, как мышечная и нервная.
Возбудимость — свойство нервной и мышечной клетки отвечать на раздражение возбуждением.
Для перехода мышцы или нерва из состояния покоя в состояние возбуждения необходимо, чтобы сила действующего раздражителя достигла критической, пороговой величины.
Чем большая возбудимость ткани, тем меньше у нее порог возбудимости. Величина порога возбудимости ткани непостоянна и зависит от ее физиологического состояния.
Для возникновения возбуждения ткани необходимо, чтобы пороговый раздражитель действовал определенное время. Наименьшее время, в течение которого должен действовать раздражитель пороговой силы, чтобы вызвать возбуждение ткани называется полезным временем. Чем сильнее раздражитель, тем короче будет время его действия, чтобы возникло возбуждение.
Если по оси абсцисс отложить время действия постоянного тока, а по оси ординат — его силу (или напряжение), то соотношение силы и длительности раздражения выразится в форме кривой, получившей название «кривая силы —времени» (рис. 74).
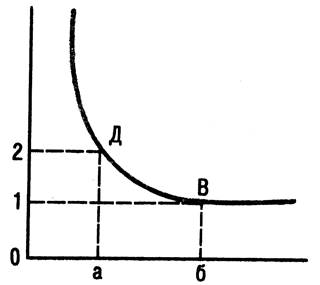
Рис. 74. Кривая силы-длительности:
0-а –хронаксия; 0-б- полезное время; 0-1- реобаза; 0-2- двойная реобаза.
Точка В, обозначающая полное время, лежит на участке кривой, идущей параллельно на оси абсцисс. Поэтому даже при очень высокой чувствительности приборов трудно точно определить полное время, так как незначительным изменениям силы (по оси ординат) будут соответствовать больше изменения во времени (по оси абсцисс). Поэтому в электрофизиологии для характеристики возбудимости по времени действия раздражителя принято брать время действия удвоенной пороговой силы этого раздражителя. Тогда точка Д, соответствующая времени действия удвоенной пороговой силы будет находиться в месте крутого изгиба кривой; в этом случае продолжительность раздражения — электрического тока — называют реобазой, а наименьшее время удвоенной реобазы — хронаксией. Хронаксия измеряется в тысячных долях секунды (сигмах) специальными приборами — хронаксиметрами.
Хронаксия зависит от структуры ткани, ее состояния, от состояния органа и всего организма.
Величина хронаксии двигательных нервов меньше, чем скелетных мышц. Например, у лошади и жвачных хронаксия двигательных нервов колеблется от 0,09 до 0,2 мс, а скелетных мышц — от 0,2 до 0,4 мс; хронаксия нервов вегетативной нервной системы — до 5 мс. Самая большая хронаксия, измеряемая сотыми и десятыми долями секунды, наблюдается у гладких мышц желудка, кишечника и матки.
Функциональная подвижность (лабильность) — одно из свойств мышечной и нервной ткани. Это свойство было открыто Н.Е. Введенским в 1897 г. при изучении действия ритмических раздражителей различной частоты на нервно-мышечный препарат.
Лабильность — это время в течение которого возникает и полностью заканчивается одиночный импульс возбуждения.
В каждой ткани одиночный импульс возбуждения продолжается определенное время. Для измерения лабильности введен показатель — мера лабильности.
Мера лабильности — максимальное число импульсов возбуждения, которые возникают за 1 с в ответ на такое же максимальное количество раздражений. Чем короче период рефрактерности, тем большее число импульсов пройдет через ткань. Наибольшей лабильностью обладают мякотные соматические нервы (500 импульсов в 1 с), для вегетативных волокон — 200 импульсов в 1 с. Для скелетных мышц — 200 импульсов в 1 с, для гладких — 10–20 импульсов в 1 с. Лабильность меняется в связи с деятельностью ткани, т.к. при начале действия раздражителя импульсы возбуждения увеличивают скорость обмена веществ в ткани, которая начинает затем воспроизводить более частый, ранее невоспроизводимый его ритм.
Изменение лабильности в сторону повышения или понижения по сравнению с исходным уровнем в связи с деятельностью ткани называется усвоением ритма. Лучше усваивается частый ритм при невысокой исходной лабильности, поэтому мышечная ткань, имеющая невысокую лабильность, обладает большей способностью к усвоению ритма, чем нервная.
Возбудимые ткани. Общие свойства. Законы раздражения.
Понятие о возбудимых тканях. Свойства живых и возбудимых тканей: раздражимость, возбудимость, проводимость, лабильность, их количественные характеристики: порог раздражения, скорость проведения возбуждения, предельный ритм раздражения. Законы раздражения: закон силы и правило «все или ничего», закон соотношения силы и длительности действия раздражителя, закон градиента.
К возбудимым тканям относятся ткани, клетки которых способны отвечать на действие раздражителя развитием потенциала действия или возбуждением. К возбудимым тканям относятся нервная, все виды мышечной ткани: скелетная, сердечная, гладкая; и железистая ткань. Этим тканям присущи общие свойства:
Раздражимость – способность ткани отвечать на действие раздражителя любым образом: изменением формы (например при сокращении мышц), интенсивности обмена веществ, проницаемости, и др.
Возбудимость– это частное проявление свойства раздражимости. Способность клетки на действие раздражителя отвечать возбуждением, т. е. развитием потенциала действия. Возбудимость у разных клеток может быть различной. Оценивается онапорогом раздражения, т. е. минимальной силой раздражителя, способного вызывать возбуждение. Как уже отмечалось выше, для развития возбуждения или потенциала действия необходимы определенные условия: пороговый раздражитель, действуя на клетку, должен снизить мембранный потенциал до критического уровня деполяризации, после чего откроются потенциалзависимые натриевые каналы и сформируется потенциал действия. Если раздражитель слабый, т. е. имеет величину меньше порогового значения, то мембранный потенциал не достигает критического уровня и возникает лишь локальный или местный ответ.
Оценивая порог раздражения можно сравнить возбудимость различных объектов. Известно, что возбудимость нервных клеток значительно выше мышечных и секреторных.
Проводимость – способность ткани проводить возбуждение. Это важное свойство нервной ткани. Отростки нейронов являются проводниками возбуждения. Показателем проводимости является скорость проведения возбуждения, она самая высокая у нервных клеток.
Лабильность – свойство ткани отвечать наибольшим числом возбуждений в единицу времени в соответствие с предложенным ритмом раздражения. Ткани, быстро реагирующие на раздражители, могут ответить в единицу времени большим числом возбуждений. Показателем лабильности является максимальное число возбуждений в единицу времени. Например, нервная клетка способна формировать до 1000 потенциалов действия в секунду.
Все названные свойства определяются свойствами плазматической мембраны.
Раздражитель – любой фактор, способный при определенных условиях вызывать возбуждение клетки.
1) физические (механические, электрические, световые, звуковые и др.);
2) химические (вкусовые, обонятельные, кислоты, газы, и др.);
3) семантические (информационные).
1) подпороговые (слабые, не способные вызывать возбуждение, но вызывающие развитие местного или локального потенциала);
2) пороговые (имеющие минимальную силу, способную вызвать
3) надпороговые (их сила больше пороговой).
с биологической точки зрения:
1) адекватные (ткань в процессе эволюции к ним приспособилась, например, свет для рецепторов зрения).
2) неадекватные раздражители (способны вызывать возбуждение только при большой силе).
Электрический ток – это универсальный раздражитель. Он широко используется в экспериментах, т. к. его легко задавать по силе, продолжительности и частоте действия.
Законы раздражения определяют, при каких условиях может возникать возбуждение.
Закон силыилизакон «всё или ничего»гласит — чтобы возникло возбуждение клетки, раздражитель должен иметь силу не менее порогового значения. Этот закон рассматривает также зависимость величины ответа от силы раздражителя. Если речь идет об отдельной клетке (нейрон, мышечное волокно), то зависимость носит название правила «все или ничего». Суть которого в следующем, если раздражитель имеет силу ниже порогового значения, ответа со стороны клетки нет — «ничего». Если раздражитель имеет силу выше порогового значения, то ответ — величина потенциала действия, остается постоянным, независимо от того, насколько раздражитель превысит пороговое значение – «всё». Если речь идет о ткани, т. е совокупности клеток, например, мышца – совокупность мышечных волокон, или нервный ствол – совокупность аксонов, то в этом случае величина ответа может быть (до определенного момента) тем больше, чем сильнее раздражитель. Это объясняется различной возбудимостью клеток. Наиболее возбудимые клетки реагируют на раздражитель меньшей силы, а по мере увеличения силы раздражителя в процесс развития возбуждения вовлекаются клетки, у которых порог раздражения выше.
Читайте также: Ткани органы системы органов анатомия
Закон соотношения силы и длительности действия раздражителя или Закон времени утверждает, что раздражитель для того, чтобы вызвать возбуждение должен быть не только достаточно сильным, но и должен действовать на ткань некоторое время, то есть быть достаточно длительным. Оказалось, что в определенном диапазоне существует обратная зависимость между силой и длительностью действия раздражителя: чем меньше времени действует раздражитель, тем выше должна быть его сила, чтобы возникло возбуждение.
Есть следствие этого закона: даже сверхсильный раздражитель, но действующий короткое время, не способен вызвать возбуждение.
Закон градиентаутверждает, что раздражитель способен вызывать возбуждение, только если он достигает порогового значения достаточно быстро. Если раздражитель имеет низкую скорость нарастания, то, чтобы вызвать возбуждение его пороговая сила должна увеличиваться. При слишком медленном нарастании силы раздражителя ткань может разрушиться, но так и не ответить возбуждением, даже если при этом раздражитель достигнет величины значительно большей порогового значения.
Дело в том, что в ответ на действие раздражителя в клетке запускаются два процесса: активирующий натриевую проницаемость мембраны за счет открытия натриевых каналов и инактивирующий натриевую проницаемость за счет закрытия натриевых каналов. При быстром нарастании силы раздражителя процесс активации опережает процесс инактивации и потенциал действия развиваются. А при медленном нарастании раздражителя инактивационные процессы могут опередить активационные и возбуждение не развивается.
Этот закон лежит в основе закаливания. Постепенно нарастающий по силе раздражитель укрепляет здоровье, не нанося вреда организму.
В целом законы раздражения определяют каким должен быть раздражитель, чтобы вызвать возбуждение: достаточно сильным, длительным и быстрым.
19. При действии подпорогового раздражителя возникает:
20. При действии сверхпорогового раздражителя возникает:
21. По силе не различают раздражители:
22. При действии порогового раздражителя возникает:
а) изменение ионной проницаемости клеточной мембраны.
б) увеличение объема цитоплазмы клетки.
в) способность переходить из состояния покоя в состояние возбуждения.
г) увеличение заряда на клеточной мембране.
д) уменьшение заряда на клеточной мембране.
а) изменение ионной проницаемости клеточной мембраны.
б) увеличение площади клеточной мембраны.
в) переход из состояния покоя в состояние возбуждения.
г) увеличение заряда на клеточной мембране.
д) способность клеточной мембраны распространять возбуждение.
а) способность с наибольшей частотой отвечать на действие раздражителя.
б) скорость течения обменных процессов в клетке.
в) способность переходить из состояния возбуждения в состояние покоя.
г) увеличение заряда на клеточной мембране.
д) способность распространять возбуждение по клеточной мембране.
26. Мерой лабильности является:
а) предельное число возбуждений.
б) скорость проведения возбуждения.
г) разность потенциалов на мембране.
27. Мерой возбудимости является:
а) предельный ритм возбуждения.
б) скорость проведения возбуждения.
г) разность потенциалов на мембране.
д) предельный ритм раздражения.
28. Мерой проводимости является
а) предельная частота возбуждений.
б) скорость проведения возбуждения.
г) разность потенциалов на мембране.
д) предельный ритм раздражения.
29. С увеличением скорости нарастания силы порогового раздражителя порог возбудимости:
30. На увеличение силы раздражителя ткань отвечает:
г) по принципу «все или ничего»
д) снижением порога возбудимости.
31. Зависимость между силой порогового раздражителя и длительностью его воздействия характеризуется как:
32. Если клетку раздражать 200 раз в секунду, то на каждый стимул возникает ПД, но если частоту раздражения увеличить, а количество ответов будет не более, чем 200 ПД/с. Эта величина является:
33. При медленном увеличении силы раздражителя, действующего на клетку:
а) порог раздражения повышается,
б) пророг раздражения снижается
в) порог раздражения не меняется
г) проявляется закон «все или ничего»
34. Укажите наиболее лабильную клетку из имеющих предельный ритм возбуждения:
35. К возбудимым тканям не относится:
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
Основные свойства живой ткани раздражимость возбудимость лабильность
tsyganskiymedvet.ru
Главная страница
Моя работа
Категории
Владельцы сайта
Возбудимые ткани
1. Возбудимые ткани, их характеристика
2. Биоэлектрические потенциалы в возбудимых тканях
3. Основные свойства возбудимых тканей
5. Строение скелетных мышц
6. Свойства скелетных мышц
7. Сила, работа, утомление мышц
1. Возбудимые ткани, их характеристика
Раздражимость (или реактивность) – это способность органов и тканей, клеток активно отвечать на воздействие какой-либо формой деятельности – обменом, ускорением деления, выбросом секрета, движением, электрическим импульсом. Ответ на раздражение является важным свойством организма.
Возбудимость – одна из форм проявления раздражимости связанная с деятельностью специализированных клеток и тканей. Способные к возбуждению клетки – мышечные, нервные, железистые – называют возбудимыми. Для возникновения возбуждения необходимо раздражение возбудимой ткани, т.е. воздействие на живую ткань раздражителя.
Раздражитель – это агент внешней или внутренней среды организма, который действуя на клетки, ткани, органы и организм в целом вызывает возбуждение.
1. По энергетической природе:
физические – механические, температурные, электрические, лучевые;
химические – кислоты, щёлочи, яды;
биологические – животные, растения, бактерии вирусы и др.
внешние (различные формы раздражения окружающей организм материи);
внутренние (изменение химического состава крови, лимфы, тканевой, спинномозговой жидкости).
пороговые – раздражители минимальной силы (энергии), необходимой для раздражения клетки. Такая сила называется пороговой (порогом). Чем ниже порог возбуждения, тем выше возбудимость (у нерва порог возбуждения ниже, чем у мышцы и особенно железы);
подпороговые – раздражители, сила которых меньше пороговой, называются;
сверхпороговые – раздражители, более сильные, чем пороговые.
4. По биологическому значению для ткани, организма:
адекватные – естественные раздражители, действующие на данную ткань в природных условиях;
неадекватные – раздражители, действию которых ткань в естественных условиях не подвергается. К ним относятся физические, механические, электрические раздражители.
При отсутствии достаточных раздражителей в клетке наблюдается состояние физиологического покоя при условии текущего метаболизма. Реакции клетки под воздействием внешних факторов или других клеток организма, выражающиеся в снижении метаболизма, роста, возбудимости, называются торможением.
Возбуждение – реакция возбудимой клетки на действие раздражителя, проявляющаяся в совокупности физических, физико-химических и функциональных изменений. Различают общие и специфические признаки возбуждения:
общие признаки – изменение мембранного потенциала (генерация потенциала действия);
специфические признаки – переход от состояния физиологического покоя к свойственной деятельности: для нервной ткани – это генерация распространяющегося импульса, синтез и разрушение медиаторов; для мышечной – сокращение; для железистой – секреция.
Для возникновения возбуждения наряду с силой и временем действия раздражителя имеет значение быстрота нарастания силы раздражителя. Так, при медленном сдавливании пинцетом нерва возбуждение не наступает, а быстром ударе по нерву моментально возникает ответная реакция, т.к. возникло возбуждение. Скорость нарастания силы раздражителя называется градиентом раздражения. Высокий градиент раздражения характеризует мгновенное нарастающее по силе раздражение до своей пороговой величины. Чем ниже градиент возбуждения, тем ниже возбудимость раздражаемых тканей. Приспособление ткани к медленно нарастающей силе раздражения называют аккомодацией. Это объясняется тем, что за длительное время действия раздражителя в клетке происходят изменения, повышающие порог возбудимости. У разных видов тканей скорость аккомодации неодинакова: высокая у двигательных нервов, мала у сердечной мышцы, гладких мышц желудка, кишечника.
Читайте также: Основные элементы хрящевой ткани
2. Биоэлектрические потенциалы в возбудимых тканях
Первые сведения о способности живых тканей генерировать электричество были получены во второй половине Х V III в. на примере рыб, имеющих электрические органы. Существование же истинного «животного электричества» как проявления процессов жизнедеятельности тканей было установлено итальянцем Л. Гальвани, опубликовавшим в 1971 г. «Трактат о силах электричества при мышечном движении». В опытах на нервно-мышечном препарате лягушки ученый обнаружил, что при замыкании между нервом и мышцей цепи из двух металлических проводников происходит сокращение мышц, что толковалось как проявление разряда электричества «предсуществовавшего» в живой ткани. Однако физик А. Вольта объяснил это явление как результат возникновения постоянного тока в цепи из двух разнородных металлов (железо, медь), где препарат служит проводником-электролитом. В 1794 г. Л. Гальвани предложил новый вариант опыта без металлических проводников: надрезал мышцу и набрасывал нерв стеклянными крючками таким образом, чтобы часть его касалась повреждённого участка, и другая часть – неповреждённого. В момент набрасывания нерва мышца сокращалась. Как доказал позже Э. Дюбуа-Реймон, мышца сокращалась из-за электрического тока, источником которого была разность потенциалов между повреждённым и неповреждённым участком мышцы (повреждённый участок заряжен отрицательно, целый – положительно. К концу ХIХ и в начале ХХ стало известно, что электрические потенциалы образуются во всех живых тканях, а возникновение и распространение возбуждения связано с изменением электрического заряда на поверхности клеточной мембраны и внутри клетки.
В настоящее время для исследования электрических явлений клетки применяют микроэлектроды (стеклянные пипетки с тонким кончиком – 0,5 мкм), заполненные электролитом. Роль проводника электрического тока играет электролит, а изолятора – стекло. Вводя кончик электрода, присоедин ё нного к регистрирующей установке, в клетку, регистрируют е ё потенциал. Благодаря данной методике в конце 50-х годов ХХ столетия А. Ходжкин, А. Хаксли, Б. Кац ( A . Hodgkin , A . Huxley , В. Katz , Великобритания) сформулировали современную мембранную теорию возникновения биоэлектрических потенциалов, получившую название теории натрий-калиевого насоса. В создание теории свой вклад также внесли В.Ю. Чаговец (Россия, 1896) и Ю. Бернштейн (Россия, 1902).
Мембрана клетки состоит из двойного слоя фосфолипидов, интегральных, трансмембранных и периферических белков, а также поверхностных гликолипидов и гликопротеинов.
В мембране существуют каналы, проницаемые для воды, для малых молекул водорастворимых веществ и для малых ионов. Особое значение имеют каналы, проницаемые для ионов Nа + , К + , Cl – , Са 2+ . В мембране обнаружены специфические (селективные) натриевые, калиевые, хлорные и кальциевые каналы, то есть каналы избирательно пропускающие названные ионы. Эти каналы могут быть открытыми или закрытыми, то есть обладают воротными механизмами. В покое практически все натриевые каналы мембраны клетки закрыты, а большинство калиевых каналов – открыты. Различная концентрация ионов по обе стороны мембраны всех клеток приводит к появлению трансмембранной разности электрического потенциала – мембранного потенциала.
Мембранный потенциал покоя
Мембранный потенциал (МП) покоя – разность электрических потенциалов между внутренней и наружной поверхностью мембраны в состоянии покоя, т.е. при отсутствии электрического или химического раздражителя (сигнала).
Потенциал покоя у разных клеток имеет величину от 50 до 90 милливольт (мВ).
Согласно современной мембранной теории потенциал покоя возникает за счет пассивного и активного движения ионов через мембрану.
Пассивное движение ионов осуществляется по градиенту концентрации и не требует затрат энергии. Клеточная мембрана в состоянии покоя более проницаема для ионов калия. Ионов калия в цитоплазме примерно в 30 раз больше, чем во внеклеточной жидкости. Они находятся в свободном состоянии и по градиенту концентрации диффундируют через клеточную мембрану во внеклеточную жидкость, в ней они не рассеиваются, а удерживаются на внешней поверхности мембраны внутриклеточными анионами.
Внутри клетки содержатся в основном анионы органических кислот: аспарагиновой, уксусной, пировиноградной и др. Содержание неорганических анионов в клетке сравнительно небольшое. Анионы не могут проникать через мембрану и остаются в клетке, располагаясь на внутренней поверхности мембраны. Так как ионы калия имеют положительный заряд, а анионы – отрицательный, то внешняя поверхность мембраны при этом заряжается положительно, а внутренняя – отрицательно.
Ионов натрия приблизительно в 10 раз больше во внеклеточной жидкости, чем в клетке, проницаемость их через мембрану незначительна. В результате пассивного движения ионов натрия и калия по концентрационному градиенту должен был бы наступить момент, когда их концентрации в клетке и внеклеточной жидкости выровнялись и разность потенциалов исчезла. Однако этого не происходит, так как в мембране существует специальный молекулярный механизм, который обеспечивает активный перенос ионов натрия и калия в сторону их повышенной концентрации и поддерживает ионную асимметрию.
Активный перенос ионов осуществляется в результате деятельности ионного Na + ,K + -насоса. Работа этого насоса заключается в перемещении ионов натрия и калия против градиента концентрации (в сторону повышенной концентрации). На каждые три иона натрия выводимые из клетки, в клетку поступают два иона калия. Na + ,K + -насос выполняет двоякую функцию: во-первых он создает и поддерживает трансмембранный градиент концентраций натрия и калия, и, во-вторых, генерирует разность потенциалов, суммирующуюся с потенциалом покоя, создаваемым пассивным движением ионов через мембрану.
Для работы насоса необходима энергия макроэргических связей АТФ. Расщепление АТФ происходит под влиянием фермента аденозинтрифосфотазы. Активность этого фермента угнетают протоплазматические яды, например, при действии цианидов прекращается работа Na + ,K + -насоса, в результате концентрация ионов по обе стороны мембраны выравнивается и клетка гибнет.
Все клетки возбудимых тканей (нервные, скелетных мышц, гладкомышечные клетки, кардиомиоциты) при действии различных раздражителей пороговой силы способны переходить в состояние возбуждения. Обязательный признак возбуждения – изменение электрического состояния клеточной мембраны. Под влиянием раздражения заряд мембраны начинает быстро уменьшаться от нуля и затем вновь появляется, но только с обратным знаком. Происходит перезарядка, или реверсия, заряда мембраны: внутренняя ее сторона заряжается положительно, а наружная – отрицательно. Затем вновь восстанавливаются первоначальные заряды мембраны. Пикообразное колебание потенциала, возникающее в результате кратковременной перезарядки мембраны и последующего восстановления ее исходного заряда называется потенциалом действия. Такое колебание потенциала продолжается 1-5 миллисекунд (мс). Величина потенциала действия нерва и скелетной мышцы составляет 115-120 мВ. В потенциале действия различают пик длительностью 0,5-0,8 мс и следовые потенциалы.
Пик потенциала действия связан с процессом возбуждения, он имеет восходящую и нисходящую фазы. Во время восходящей фазы вначале исчезает исходная поляризация мембраны, поэтому её называют фазой деполяризации, затем изменяется знак потенциала – реверсия. Во время нисходящей фазы заряд мембраны возвращается к уровню покоя, и её называют реполяризацией. При реполяризации отмечают следовые потенциалы: следовую деполяризацию и следовую гиперполяризацию. Следовые потенциалы связаны с восстановительными процессами после окончания возбуждения.
Читайте также: Цветы из ткани инструменты для изготовления цветов
Современная мембранная теория объясняет механизм возникновения потенциала действия следующим образом. При действии раздражителей пороговой или сверхпороговой силы наступает деполяризация клеточной мембраны. Когда деполяризация достигает некоторой критической величины, одновременно за доли миллисекунды открывается максимальное количество натриевых каналов, проницаемость мембраны для ионов натрия возрастает в 500 раз по сравнению с покоем. В результате этого ионы натрия из внеклеточной жидкости, где их концентрация повышена, лавинообразно устремляются внутрь клетки и уносят с наружной поверхности мембраны положительные заряды, значительно увеличивая их концентрацию в клетке; внутренняя поверхность мембраны приобретает положительный заряд. Вследствие уменьшения положительных зарядов наружная поверхность мембраны за счет внеклеточных анионов заряжается отрицательно. Но увеличение натриевой проницаемости очень кратковременно и длится примерно 0,5-1,5 мс, после чего проницаемость мембраны для ионов натрия вновь понижается до исходного уровня в результате уменьшения количества открытых натриевых каналов. Вслед за этим проницаемость мембраны для ионов калия повышается в 10-15 раз. Они начинают усиленно выходить из клетки и, унося из нее положительные заряды, восстанавливают исходную разность потенциалов, то есть мембрана опять приобретает снаружи положительный заряд, а изнутри – отрицательный.
Следовательно, потенциал действия возникает в результате прохождения ионных потоков через мембрану: движение ионов натрия внутрь клетки приводит к перезарядке мембраны, а выход из клетки ионов калия восстанавливает исходный потенциал покоя. Потоки приблизительно равны по величине, но сдвинуты во времени.
3. Основные свойства возбудимых тканей
Для возбудимых клеток и тканей характерны раздражимость, возбудимость, проводимость, функциональная подвижность (лабильность).
1. Раздражимость – общее свойство любой живой ткани, клетки реагировать на раздражение изменением обмена веществ и энергии. Следует учитывать, что изменения носят местный характер, т.е. реагирует только участок ткани, который подвергается раздражению.
2. Возбудимость – это свойство нервной, мышечной или железистой клетки отвечать на действие раздражителей изменениями ионной проницаемости мембраны и генерировать потенциал действия. Для возникновения состояния возбуждения необходимы определенные условия:
а) сила действующего раздражителя должна быть пороговой;
б) достаточное время действия раздражителя (полезное время). Чем сильнее раздражитель, тем меньше времени потребуется для возбуждения ткани.
в) действие раздражителя с большой силой нарастания (градиентом).
Для изучения возбуждения, как формы раздражимости, у живых организмов электрический ток имеет преимущество, так как легко дозируется по силе, деятельности, характеру, вызывает возбуждение, не повреждая, и после прекращения раздражения не оставляет необратимых изменений (ток образуется и в живых тканях, поэтому действие электрического тока близко к природным механизмам).
Для характеристики возбудимости тканей в электрофизиологии приняты следующие термины:
реобаза – пороговая сила раздражителя – электрического тока;
полезное время – время, необходимое для того, чтобы раздражитель пороговой силы вызвал возбуждение;
хронаксия – наименьшее время действия удвоенной реобазы (измеряют в миллисекундах или сигмах). Чем меньше хронаксия, т.е. сила тока, способная вызвать возбуждение, тем выше возбудимость ткани. Хронаксия – величина переменная, зависящая от структуры ткани, её физиологического состояния. У скелетных мышц величина хронаксии больше, чем у двигательных нервов. Самая большая хронаксия, измеряемая сотыми и десятыми долями секунды, – у гладких мышц кишечника, желудка, матки.
Изменение возбудимости ткани при возбуждении.
Для возбудимости тканей характерны фазовые изменения в зависимости от возникновения и развития импульсов. Во время деполяризации мембраны, при действии возбудителя, и появлении местного возбуждения происходит кратковременное повышение возбудимости. Когда местное возбуждение переходит в распространяющееся, возникает потенциал действия, а возбудимость ткани резко снижается. Возникает состояние, когда ткань после раздражения временно не реагирует на повторное раздражение любой силы – абсолютная рефрактерность (совпадает с пиком потенциала действия) – от лат. refrakta – невосприимчивость. Длительность фазы – в нервных волокнах – 1-10, в мышцах – до 30 мс. Период восстановления ткани после абсолютной рефрактерности, когда возбудимость ещё понижена называется относительной рефрактерностью (совпадает с фазой реполяризации и переходом его в следовую поляризацию). Длительность периода – в нерве – 20, в мышце – 50 мс.
Затем наступает следующий период – фаза экзальтации (от лат. exsaltatio – очень возбужденный) – период повышенной возбудимости, совпадающий с периодом следовой поляризации. Длительность – в нерве – 20, в мышцах – 50 мс. Со следовой гиперполяризацией совпадает фаза субнормальности, во время которой возбудимость ткани несколько снижена по сравнению с величиной возбудимости в состоянии физиологического покоя.
Очевидно, что фазы изменения возбудимости определены развитием и изменением потенциала действия в живой ткани. Изменение возбудимости зависит от действия натрий-калиевого насоса.
Самый частый ритм раздражений, на которые возбудимая ткань отвечает таким же частым ритмом возбуждения, отражает функциональное состояние ткани или лабильность. В 1928 году А.А. Ухтомский создал представление об усвоении ритма, согласно которому лабильность может изменяться под действием разных условий, в частности зависеть от обмена веществ в ткани. Понижение и повышение физиологической лабильности в связи с деятельностью, проявляющееся более высоким или низким ритмом возбуждения названо ученым усвоением ритма.
При изучении влияния на живую ткань частоты ритмов раздражения Н.Е. Введенский установил ритмы, наиболее выгодные в рабочем отношении, так называемый оптимум. При этом ритме раздражений новый импульс возникает в момент фазы экзальтации предшествующего импульса, в результате мышца сокращается максимально. Показатели оптимального ритма меньше максимального, Так максимальный ритм для двигательного нерва лягушки – 500, а оптимальный 75-150 импульсов, для мышечных волокон максимальный 150 импульсов/с, оптимальный – 20 – 50 в 1 с. Пессимум ритма раздражения возникает, когда частота раздражений настолько высока, что превышает меру лабильности. Это вызвано тем, что раздражение действует в фазу абсолютной рефрактерности предыдущего импульса. Фаза рефрактерности может углубиться, лабильность будет продолжать снижаться и наступит торможение. Перечисленные явления характерны не только для двигательных реакций, но и для условно-рефлекторных, для секреторных реакций, деятельности сердечно-сосудистой системы.
Н.Е. Введенский в классическом труде «Возбуждение, торможение и наркоз» изложил каким образом возбуждение переходит в торможение. Ученый воздействовал на среднюю часть нерва нервно-мышечного препарата лягушки эфиром, теплом, холодом, хлористым калием, сильным электрическим током, т.е. вызывал альтерацию ткани. По обеим сторонам участка накладывал электроды индукционной катушки. Электрическую активность изучал при помощи гальванометра. Ответная реакция мышцы на раздражения была неоднозначной, что дало основание выделить несколько последующих стадий:
1 стадия – уравнительная или трансформирующая. При действии перечисленных веществ различной природы, отличающихся по силе и частоте раздражения, мышца начинает отвечать одинаковыми сокращениями;
2 стадия – парадоксальная, сопровождалась сильными сокращениями мышцы при слабых или редких раздражениях, а при действии сильных раздражителей, наоборот, не сокращалась или почти не реагировала;
3 стадия – парабиотическая, сопровождалась состоянием, когда при воздействии на нерв раздражителя любой силы и частоты мышца не сокращалась, развивалось торможение, заканчивающееся состоянием, при котором возбудимость и проводимость, как проявления жизни, отсутствуют.
Эти последовательные функциональные изменения были названы Введенским – стадиями парабиотического процесса. При удалении раздражающих веществ с нерва парабиоз прекращается, функциональные свойства участка альтерируемой ткани восстанавливаются, проходя те же стадии в обратном порядке. Если воздействие раздражителей длительное, нерв отмирает.
5. Строение скелетных мышц
У всех высших животных мышцы являются важнейшими исполнительными, т.е. рабочими органами (эффекторами). У позвоночных различают мышцы: