
48. Особенности обмена глюкозы в разных органах и клетках: эритроциты, мозг, мышцы, жировая ткань, печень.
Метаболизм глюкозы в эритроцитах. Эритроциты лишены митохондрий, поэтому в качестве энергетического материала они могут использовать только глюкозу. В эритроцитах катаболизм глюкозы обеспечивает сохранение структуры и функции гемоглобина, целостность мембран и образование энергии для работы ионных насосов. Глюкоза поступает в эритроциты путём облегчённой диффузии с помощью ГЛЮТ-2. Около 90% поступающей глюкозы используется в анаэробном гликолизе, а остальные 10% — в пентозофосфатном пути. Конечный продукт анаэробного гликолиза лактат выходит в плазму крови и используется в других клетках, прежде всего гепатоцитах. АТФ, образующийся в анаэробном гликолизе, обеспечивает работу Nа + , К+-АТФ-азы и поддержание самого гликолиза, требующего затраты АТФ в гексокиназной и фосфофруктокиназной реакциях. Важная особенность анаэробного гликолиза в эритроцитах по сравнению с другими клетками — присутствие в них фермента бисфосфоглицератмутазы. Бисфосфоглицератмутаза катализирует образование 2,3-бисфосфоглицерата из 1,3-бисфосфоглицерата. Образующийся только в эритроцитах 2,3-бисфосфоглицерат служит важным аллостерическим регулятором связывания кислорода гемоглобином. Глюкоза в эритроцитах используется и в пентозофосфатном пути, окислительный этап которого обеспечивает образование кофермента NADPH, необходимого для восстановления глу-татиона.
Метаболизм глюкозы в печени.Основная рольпеченив углеводном обмене заключается в обеспечении постоянстваконцентрацииглюкозывкрови. Это достигается регуляцией междусинтезом и распадом гликогена, депонируемого впечени. Впеченисинтез гликогенаи его регуляция в основном аналогичны тем процессам, которые протекают в других органах итканях, в частности вмышечной ткани.Синтез гликогенаизглюкозыобеспечивает в норме временный резервуглеводов, необходимый для поддержанияконцентрацииглюкозывкровив тех случаях, если ее содержание значительно уменьшается (например, у человека это происходит при недостаточном поступленииуглеводовс пищей или в период ночного «голодания»). Необходимо подчеркнуть важную рольферментаглюкокиназы в процессе утилизацииглюкозыпеченью. Глюкокиназа, подобногексокиназе, катализируетфосфорилированиеглюкозыс образованием глюкозо-6-фосфата, при этомактивностьглюкокиназы впеченипочти в 10 раз превышаетактивностьгексокиназы. Важное различие между этими двумяферментамизаключается в том, что глюкокиназа в противоположностьгексокиназеимеет высокое значение КМ дляглюкозыи не ингибируется глюкозо-6-фосфатом. Считают, что основная рольпечени– расщеплениеглюкозы– сводится прежде всего к запасанию метаболитов-предшественников, необходимых длябиосинтеза жирных кислотиглицерина, и в меньшей степени кокислениюее до СО2 и Н2О. Синтезированные впеченитриглицериды в норме выделяются вкровьв составелипопротеинови транспортируются вжировую тканьдля более «постоянного» хранения. Вреакцияхпентозофосфатного пути впечениобразуется НАДФН, используемый для восстановительныхреакцийв процессах синтезажирных кислот,холестеринаи другихстероидов. Кроме того, при этом образуются пентозофосфаты, необходимые для синтезануклеиновых кислот. Наряду с утилизациейглюкозывпеченипроисходит и ее образование. Непосредственным источникомглюкозывпеченислужитгликоген.Распад гликогенавпеченипроисходит в основном фосфоролитическим путем. В регуляции скоростигликогенолизавпеченибольшое значение имеет система циклическихнуклеотидов. Кроме того,глюкозавпечениобразуется также в процессеглюконеогенеза.
Метаболизм глюкозы в мозгу.Основнымсубстратомдыханиямозговойтканиявляетсяглюкоза. В 1 мин 100 гтканимозга потребляют в среднем 5 мгглюкозы. Подсчитано, что более 90% утилизируемойглюкозывтканимозга окисляется до СО2и Н2О при участиицикла трикарбоновых кислот. В физиологических условиях роль пентозофосфатного путиокисленияглюкозыв мозговойтканиневелика, однако этот путьокисленияглюкозыприсущ всемклеткамголовного мозга. Образующаяся в процессепентозофосфатного циклавосстановленная форма НАДФ (НАДФН) используется для синтезажирных кислотистероидов. Междуглюкозойигликогеноммозговойтканиимеется тесная связь, выражающаяся в том, что при недостаточном поступленииглюкозыизкровигликогенголовного мозга является источникомглюкозы, аглюкозапри ее избытке – исходным материалом длясинтеза гликогена.Распад гликогенав мозговойтканипроисходит путем фосфоролиза с участием системыцАМФ. Однако в целом использованиегликогенав мозге по сравнению сглюкозойне играет существенной роли в энергетическом отношении, так как содержаниегликогенав головном мозге невелико. Наряду с аэробнымметаболизмом углеводовмозговаятканьспособна к довольно интенсивному анаэробномугликолизу. Значение этого явления пока недостаточно ясно, ибогликолизкак источник энергии ни в коей мере не может сравниться по эффективности с тканевымдыханиемв головном мозге.
Метаболизм глюкозы в мышцах.При работе умеренной интенсивности мышца может покрывать свои энергетические затраты за счет аэробногометаболизма. Однако при больших нагрузках, когда возможность снабжениякислородомотстает от потребности в нем, мышца вынуждена использовать гликолитический путь снабжения энергией. При интенсивной мышечной работе скорость расщеплениягликогенаилиглюкозыс образованиеммолочной кислотыувеличивается в сотни раз. Соответственно содержаниемолочной кислотывмышечной тканиможет повышаться до 1,0–1,2 г/кг и более. С токомкровизначительное количествомолочной кислотыпоступает впечень, где ресинтезируется вглюкозуигликоген(глюконеогенез) за счет энергии окислительных процессов.
Метаболизм глюкозы в жировой ткани. В адипоцитах для обеспечения реакций синтеза жира распад глюкозы идёт по двум путям: гликолиз, обеспечивающий образование глицерол-3-фосфата и ацетил-КоА, и пентозофосфатный путь, окислительные реакции которого обеспечивают образование NADPH, служащего донором водорода в реакциях синтеза жирных кислот.
Гликолиз в мышечных волокнах
Дается определение одного из анаэробных путей ресинтеза АТФ в мышечных волокнах – гликолиза. Описаны количественные критерии гликолиза в мышечных волокнах : максимальная мощность, время развертывания, время работы с максимальной мощностью. Дается характеристика ключевым ферментам гликолиза, протекающего в мышечных волокнах: фосфорилазе и фосфофруктокиназе.
Гликолиз в мышечных волокнах
Определение
Под гликолизом понимается процесс окисления глюкозы, при котором из одной молекулы глюкозы образуются две молекулы пировиноградной кислоты (Н.И.Волков с соавт. 2000).
Однако, как указывает С.С. Михайлов (2009) основной углевод мышечных волокон — гликоген. Свободная глюкоза в саркоплазме содержится в очень малой концентрации — имеются лишь ее следы. Поэтому в мышечных волокнах гликолиз протекает в виде распада гликогена. А.Дж. Мак-Комас (2001) по этому поводу пишет так: «Хотя мышечные волокна способны окислять глюкозу, поступающую из кровотока через интерстициальную жидкость, во время сокращений основное количество окисляемой глюкозы обеспечивается гидролизом гликогена«. С. 231.
Гликолиз, протекающий в мышечных волокнах (гликолитический ресинтез АТФ, лактатный ресинтез АТФ) – анаэробный распад гликогена мышц до молочной кислоты (лактата).
Источником энергии, необходимой для ресинтеза АТФ является мышечный гликоген, который находится в саркоплазме мышечных волокон. Концентрация гликогена в саркоплазме составляет 0,5-2% от массы мышцы (С.С. Михайлов, 2009). Также может быть использована глюкоза, поступающая из крови (Н.И. Волков с соавт., 2000), рис.1.
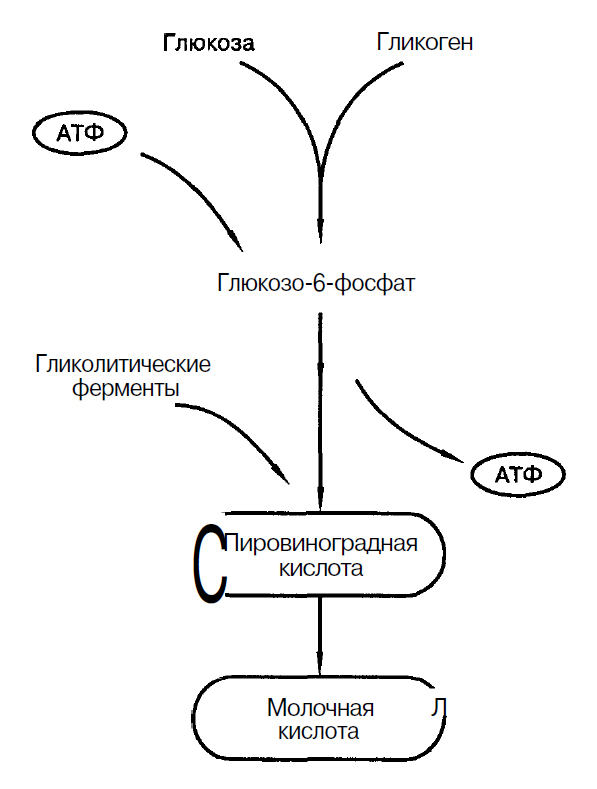
Рис. 1. Процесс гликолиза (Дж. Уилмор, Д.Л. Костилл, 1997)
Гликолиз – основной механизм энергообеспечения при работе субмаксимальной мощности длительностью 2-3 мин. Поэтому видами спорта, в которых гликолиз является основным механизмом обеспечения являются: бег на 800 – 1500 м; бег на коньках на 1500 м; плавание на 200 м и др.
Ключевыми ферментами гликолиза являются: фосфорилаза и фосфофруктокиназа. Эти ферменты регулируют скорость протекания гликолиза. Фермент фосфорилаза активируется адреналином.
Количественные критерии гликолиза в мышечных волокнах
Максимальная мощность
Максимальная мощность гликолиза составляет 750-850 кал/мин кг. Этот показатель в два раза превышает мощность тканевого дыхания. Высокие значения максимальной мощности объясняются большим количеством гликогена, содержащегося в мышечных волокнах, наличием механизмов активации ключевых ферментов гликолиза, благодаря которым скорость гликолиза возрастает в 2000 раз, отсутствием потребности в кислороде.
Время развертывания гликолиза
Время развертывания гликолиза составляет 20-30 с. Достаточно небольшое время развертывания гликолиза объясняется тем, что все участники этой реакции, а именно гликоген и ключевые ферменты находятся в саркоплазме. Кроме того, ключевой фермент гликолиза – фосфорилаза, активируется стрессовым гормоном адреналином, который выделяется в кровь непосредственно перед началом физической нагрузки. Также фермент фосфорилаза активируется ионами кальция, концентрация которых в саркоплазме повышается более чем в 1000 раз при развитии потенциала действия.
Время работы с максимальной мощностью
Время работы с максимальной мощностью гликолиза составляет всего 2-3 минуты. Это связано с тем, что, во-первых, гликолиз протекает с высокой скоростью, что приводит к быстрому исчерпанию гликогена мышц. Во-вторых, в процессе гликолиза в саркоплазме повышается концентрация молочной кислоты, которая быстро диссоциирует. В результате в мышечных волокнах накапливается лактат и протоны водорода. В-третьих, в настоящее время установлено, что основным поставщиком протонов водорода является гидролиз АТФ , а не гликолиз, как думали раньше. Эта реакция при работе субмаксимальной мощности протекает очень активно. Накопление протонов водорода в результате гидролиза АТФ и гликолиза вызывает сдвиг рН саркоплазмы в кислую сторону и снижает активность ключевых ферментов гликолиза. При значении рН саркоплазмы равном 6,4 расщепление гликогена прекращается. Это вызывает резкое снижение уровня АТФ и развитие утомления (Н.И. Волков с соавт., 2000).
Читайте также: Волокнистую структуру имеет ткань
В покое до физической нагрузки концентрация лактата в крови составляет 1-2 ммоль/л. После физической нагрузки субмаксимальной мощности концентрация лактата в крови резко возрастает и может достигать 18-20 ммоль/л.
Если физическая активность продолжается более 20 минут, запасы гликогена в мышечных волокнах истощаются. Основным источником АТФ в мышцах становится окисление жирных кислот в митохондриях мышечных волокон. Именно по этой причине считается, что для профилактики борьбы с ожирением нужны длительные циклические нагрузки аэробного характера.
Экономичность гликолиза
Гликолиз характеризуется низкой экономичностью. Распад до молочной кислоты одного остатка глюкозы дает только три молекулы АТФ, в то время как при аэробном окислении гликогена до воды и углекислого газа образуется 39 молекул АТФ в расчете на один остаток глюкозы.
Следует отметить, что гликолиз активно протекает в мышечных волокнах IIА и IIB типа.
Параграфы 32 и 34 Гликолиз. Аэробный обмен углевод
Автор текста – Анисимова Е.С.
Продавать текст нельзя. Авторские права защищены.
Для замечаний:
https://vk.com/bch_5
Параграфы 32 и 34.
Реакции, формулы и ферменты см. в файле «32 ФОРМУЛЫ ГЛИКОЛИЗа».
ПАРАГРАФ 32:
«Гликолиз».
Параграф 34:
«Аэробный обмен углеводов».
Гликолиз – это процесс превращения глюкозы в пируват (10 реакций).
Считается вторым этапом катаболизма (второй называется унификацией) в обмене углеводов – см. п.20.
Локализация процесса – гликолиз протекает во всех клетках организма, в гиалоплазме.
Глюкоза для гликолиза и других процессов поступает в клетки из крови, для чего в крови должна поддерживаться достаточная концентрация глюкозы (от 3 ммоль/л – см. п.37).
Транспорт глюкозы через мембраны клеток осуществляется специальными белками-переносчиками глюкозы. Транспорт глюкозы в клетки многих тканей (например, жировой и мышечной тканей, но не нейронов и эритроцитов) стимулируется инсулином (см. п.102, 103); и поэтому при снижении количества инсулина в крови или снижении действия инсулина (при диабете) транспорт глюкозы в клетки названных тканей снижен, что приводит к дефициту глюкозы в клетках и к избытку глюкозы в крови – см. п.37.
Кроме того, при расщеплении гликогена в мышцах (гликогенолизе – см.п.31) образуется метаболит гликолиза – глюкозо-6-фосфат, который сразу вступает во вторую реакцию гликолиза.
32.2. Конечные метаболиты гликолиза, значение и тканевые особенности
(то есть в каких клетках и тканях какие варианты бывают).
Центральным конечным метаболитом гликолиза является ПИРУВАТ (2 молекулы пирувата образуются из одной молекулы глюкозы).
Дальнейшие превращения пирувата зависят от условий, в которых протекает гликолиз – или в лактат, или в ацетилКоА, или в оксалоацетат (см. далее).
32.2.1. Превращения пирувата В АЭРОБНЫХ УСЛОВИЯХ
(АЭРОБНЫЙ ОБМЕН УГЛЕВОДОВ:
это гликолиз до пирувата и дальнейшие превращения пирувата, о которых тут сказано):
При наличии кислорода и возможности его использования (то есть в аэробных условиях в митохондриях во всех клетках, кроме эритроцитов)
1 – пируват может превратиться в АЦЕТИЛ-КоА под действием ПДГ (см. п.20, 21, «32 формулы гликолиз»), что даёт по 2,5 АТФ от каждой молекулы пирувата.
Пируват ; ацетилКоА + 2,5 АТФ
2.1 – АцетилКоА может поступить в ЦТК, что может привести к выработке 10 молекул АТФ от каждой молекулы ацетилКоА. И превращению ацетилКоА в углекислый газ для выдоха. Что завершает распад углеводородного скелета молекулы глюкозы.
ацетилКоА ; 2СО2 + 10 АТФ (при поступлении в ЦТК и ДЦ)
2.2 – В печени ацетилКоА может использоваться для синтеза жирных кислот (при сытости, см. п.46),кетоновых тел (при голоде и стрессе, см. п.47) и холестерина (см. п.49), а также (через реакции ЦТК) – аминокислот глутамата, глутамина и т.д. (см. п.67).
ацетилКоА ; жирные кислоты, кетоновые тела, холестерин, аминокислоты (с оксалоацетатом)
3 – Из пирувата может образоваться аминокислота АЛАНИН (см. п.64, 67 и 33) и кетокислота ОКСАЛОАЦЕТАТ (см. п.33), которая может 1) использоваться в ЦТК (см.п. 21) и 2) для синтеза аминокислоты аспартата для синтеза аспарагина, белков, для обезвреживания аммиака в синтезе мочевины (см.п.66) и для синтеза нуклеотидов (а затем РНК и ДНК).
4 – из метаболита гликолиза 3-фосфоглицерата (см. п.67) может синтезироваться аминокислота СЕРИН, из которой могут синтезироваться глицин, цистеин, липоиды (см. п.52).
Пируват ; аланин или оксалоацетат (; аспартат ; мочевина или нуклеотиды ; ДНК и РНК)
ОКСАЛОАЦЕТАТ для ЦТК или ; аспартат ; мочевина или нуклеотиды ; ДНК и РНК)
; (биотин!)
Аланин ; ПИРУВАТ ; (ПДГ) АЦЕТИЛ-КоА (+ 2,5 АТФ) ; (в ЦТК) 2СО2 + 10 АТФ (всего 12,5 АТФ)
; (под действием ПДГ, витамины – В1, В2, РР, пантотенат, липоевая)
ацетилКоА ; жирные кислоты (для жира и липоидов), кетоновые тела, холестерин
ЗНАЧЕНИЕ ГЛИКОЛИЗА в аэробных условиях – в получении всех перечисленных веществ, в том числе для синтезов (можно сказать, что это анаболическая роль гликолиза по аналогии с анаэробной ролью ЦТК). Участие гликолиза в выработке АТФ – это энергетическая роль гликолиза. Участие гликолиза в катаболизме глюкозы (превращение 6-тиуглеродной молекулы глюкозы в две трёхуглеродных молекулы пирувата) и поставка метаболитов для ЦТК (ацетилКоА и оксалоацетата) – катаболическая. Это достоинства гликолиза в аэробных условиях.
Недостатком аэробного обмена глюкозы является то, что его реакции протекают только в аэробных условиях и не протекают в анаэробных.
32.2.2. Превращения пирувата В АНАЭРОБНЫХ УСЛОВИЯХ.
При отсутствии кислорода или невозможности его использования в дыхательной цепи (см. п.22), то есть в анаэробных условиях пируват превращается в ЛАКТАТ (под действием фермента ЛДГ),
поскольку в анаэробных условиях пируват не может превратиться в ацетилКоА, так как не работает ПДГ (из-за избытка НАДН и дефицита НАД).
Причиной сниженной активности ПДГ и образования лактата может быть и дефицит витаминов ПДГ, особенно В1.
Лактат – это анион молочной кислоты, которая при клеточном рН (около 7) диссоциирует на лактат и протон; повышение концентрации протонов (Н+) называется ацидозом («закислением»), поскольку именно наличие протонов создаёт кислую среду, кислую реакцию, снижает рН.
Использовать кислород в дыхательной цепи не могут ЭРИТРОЦИТЫ из-за отсутствия в них митохондрий и дыхательной цепи,
а также любые клетки, если в них не работает дыхательная цепь (см.п.22) из-за попадания в них ингибиторов дыхательной цепи.
Дефицит кислорода (гипоксия) возникает во всех клетках при нарушении доставки кислорода:
1) из-за нарушения кровотока (из-за тромбов, жгута, сдавливания, падения артериального давления, большой кровопотери, сердечной недостаточности и т.д.),
2) из-за нарушения работы эритроцитов (см. п.121),
3) из-за нарушения поступления кислорода в лёгкие (при эмфиземе лёгких у курильщиков, при бронхоспазме и т.д.).
Превращение глюкозы в лактат в анаэробных условиях могут называть анаэробным гликолизом.
Лактат опасен, поскольку его накопление приводит к АЦИДОЗУ, который называется лактацидозом (в отличие от кетоацидоза при накоплении кетоновых тел – см. п. 47). Лактацидоз и кетоацидоз относятся к метаболическим ацидозам, то есть к ацидозам, обусловленным накоплением метаболитов (кислот) и нарушением метаболизма. Есть ещё респираторный ацидоз – то есть связанный с нарушением дыхания (задержкой дыхания). При всех ацидозах портится самочувствие, нарушается работа белков (так как оптимум для большинства белков – рН около 7), возникает чувство тошноты. Сильный ацидоз может привести к коме, но для здоровых людей это не характерно.
Читайте также: Клей спреем для ткани
32.3. ОБМЕН ЛАКТАТА.
Это процессы образования и использования лактата.
Как сказано выше, образуется лактат при анаэробном гликолизе в эритроцитах, мышцах и других клетках в анаэробных условиях. Из клеток он может поступать в кровь, что при избытке лактата приводит к ацидозу.
Ткани-клетки-источники лактата:
В покое основное количество лактата поступает в кровь из эритроцитов, а при наряжённой работе мышц – из мышц.
Кроме этого, лактат поступает в кровь из любых клеток, которым не хватает кислорода или в которых не работают дыхательная цепь, ЦТК и ПДГ. Причиной сниженной работы ЦТК бывает дефицит витаминов ЦТК, особенно В1. См. 32.2.2.
32.3.2. Использование и УТИЛИЗАЦИЯ ЛАКТАТА.
Из крови большая часть лактата поступает в клетки печени, где утилизируется. Это снижает ацидоз. Поэтому нарушение работы печени (печёночная недостаточность) может стать причиной избытка лактата в крови (из-за сниженного использования лактата клетками печени).
Пути метаболизма лактата – 25% и 75%.
25% лактата подвергаются так называемому ОКИСЛЕНИЮ ЛАКТАТА.
При этом:
1) лактат превращается в пируват под действием фермента ЛДГ, что даёт 2,5 АТФ благодаря образованию НАДН и его использованию в дыхательной цепи,
2) затем пируват превращается в ацетилКоА под действием ПДГ, что даёт ещё 2,5 АТФ благодаря образованию НАДН и его использованию в дыхательной цепи,
3) затем ацетилКоА вступает в ЦТК, что даёт ещё 10 АТФ.
В сумме окисление одной молекулы лактата даёт 15 молекул АТФ.
Реакции окисления лактата идут только в аэробных условиях.
75% лактата используются для синтеза глюкозы, который называется глюконеогенезом (ГНГ) и протекает в основном в печени.
Поэтому снижение ГНГ может стать причиной избытка лактата и ацидоза.
При превращении лактата в глюкозу:
1) сначала лактат превращается в пируват под действием ЛДГ, как и при окислении лактата,
2) затем пируват превращается в ОКСАЛОАЦЕТАТ (см. п.33 и п.21), что требует витамина БИОТИН – без биотина реакция не идёт (причиной дефицита биотина, нарушения этой реакции, ГНГ и избытка лактата бывает избыточное употребление сырых яиц, так как в них содержится вещество, препятствующее усвоению биотина),
3) оксалоацетат превращается в глюкозу в ходе реакций глюконеогенеза.
32.3.3. Избыток лактата.
Между процессами образования лактата и его использования должно быть равновесие, благодаря которому концентрация лактата в клетках и крови не превышала бы нормальную.
Повышение концентрации лактата в клетках приводит к снижению рН в клетках, нарушению работы белков клеток, повреждению клеток. Например, мышцы при избыточной концентрации лактата теряют способность сокращаться.
Повышение концентрации лактата в крови называется гиперЛАКТАТемией и приводит к ацидозу.
К избытку лактата приводит или повышение образования лактата, или снижение утилизация лактата.
Причины повышенного образования лактата (см. выше 32.2.3. и 32.3) –
1) дефицит кислорода (гипоксия при анемии, сердечной или лёгочной недостаточности и т.д.) или снижение его использования,
2) снижение превращения пирувата в ацетилКоА из-за нарушения работы ПДГ при дефиците В1 или других витаминов ПДГ,
3) повышенная концентрация НАДН, которая бывает не только при дефиците кислорода, но и при употреблении алкоголя.
Причины сниженного использования лактата и пирувата:
1) дефицит витаминов ПДГ и биотина, без которых не протекают реакции использования и пирувата,
2) нарушение работы печени, в котором должен протекать ГНГ.
Для коррекции ацидоза, обусловленного избытком лактата, нужно убрать причины избытка лактата – восстановить кровоток, принимать витамины, не употреблять алкоголь и т.д.
32.4. Выработка АТФ при анаэробном гликолизе
(энергетический баланс анаэробного гликолиза).
В анаэробных условиях гликолиз (при котором глюкоза превращается в лактат) позволяет синтезировать 2 молекулы АТФ способом субстратного фосфорилирования (см. файл «32 формулы гликолиз» реакции № 7 и 10, а также п.23) в расчёте на использование одной молекулы глюкозы (превращение одной глюкозы в 2 молекулы лактата).
1. Этого количества хватает только эритроцитам (у них и нет других источников АТФ).
Остальным клеткам этого (2 молекул АТФ от одной молекулы глюкозы) мало, но без анаэробного гликолиза клетки не имели бы и этого в анаэробных условиях.
2. При наложении жгута и в других ситуациях нарушения кровотока ткани конечностей не подвергаются в течение часа некрозу именно благодаря тому, что есть небольшое количество АТФ, которое вырабатывается при гликолизе в анаэробных условиях. А после часа времени возникает риск некроза именно потому, что АТФ от анаэробного гликолиза мало.
3. Клеткам головного мозга недостаточно того количества АТФ, который может им дать анаэробный гликолиз, поэтому они не способны, в отличие от тканей конечностей, выдержать гипоксию в течение часа.
4. При интенсивной работе мышц (например, при беге на сто метров или при подъёме штанги) в них возникают анаэробные условия. Наличие в них АТФ для работы поддерживается именно благодаря 2 АТФ анаэробного гликолиза, который быстро «включается» при работе мышц.
Ранее анаэробного гликолиза АТФ образуется благодаря запасам креатин/фосфата – см. п.68. Более длительная работа мышц требует получения АТФ при аэробных процессах – аэробном обмене глюкозы и при ;-окислении жирных кислот.
32.5. Достоинства и недостатки анаэробного гликолиза.
Таким образом, то, что анаэробный гликолиз даёт хоть немного АТФ без кислорода – это его ДОСТОИНСТВО.
А то, что этого количества АТФ МАЛО (хватает только эритроцитам, тканям конечностей при гипоксии и скелетным мышцам при короткой напряжённой работе, но не хватает другим тканям вообще, а тканям конечностей не хватает более чем на час, не хватает мышцам для длительной работы – это НЕДОСТАТОК анаэробного гликолиза).
Ещё один недостаток анаэробного гликолиза – то, что избыток его конечного метаболита (лактата) приводит к ацидозу.
Гликолиз является единственным процессом, за счёт которого может образоваться АТФ при отсутствии (или неиспользовании) кислорода. Именно гликолиз обеспечивает молекулами АТФ эритроциты, в которых нет других источников АТФ из-за отсутствия митохондрий и дыхательной цепи см. п.23.
32.6. Выработка АТФ при аэробном обмене глюкозы.
См. «32 формулы гликолиз» и 32.2.
В аэробных условиях глюкоза превращается в 6 молекул СО2, что сопровождается синтезом 32 молекул АТФ.
При гликолизе глюкоза превращается в 2 молекулы пирувата, что сопровождается образованием 2 молекул АТФ способом субстратного фосфорилирования и двух молекул НАДН (см. реакцию гликолиза № 6).
Каждая из этих 2 молекул НАДН даёт по 2,5 АТФ, то есть всего 5 (с помощью ДЦ).
Каждая из двух молекул пирувата даёт по 12,5 молекул АТФ, то есть всего – 25.
В сумме глюкоза может дать 32 молекулы АТФ. (2 + 2,5*2 + 12,5*2).
В головном мозге из-за особенностей обмена глюкоза даёт не 32, а 30 молекул АТФ.
32.7. РЕГУЛЯЦИЯ ГЛИКОЛИЗА. См. п.6 и 7, 33, 37, 102.
Смысл регуляции любого процесса в том, чтобы его скорость соответствовала потребностям организма и отдельных клеток.
Изменение скорости процесса осуществляется через изменение активности или концентрации ферментов процесса под влиянием метаболитов и гормонов.
Активность повышается активаторами и снижается ингибиторами.
Концентрация ферментов повышается в результате усиления синтеза ферментов под влиянием индукторов и снижается в результате снижения синтеза под влиянием репрессоров.
Читайте также: Выкройка юбки из джинсовой ткани для девочки
Регулировать все ферменты 10 реакций гликолиза не обязательно. Достаточно регулировать активность некоторых ферментов. Эти регулируемые ферменты называются КЛЮЧЕВЫМИ – см. п.7.
В гликолизе ключевыми ферментами являются три – ферменты первой реакции, последней и третьей – гексокиназа, фосфофруктокиназа и пируваткиназа.
Фермент первой реакции последовательности реакций часто является ключевым. Фермент третьей реакции гликолиза является самым медленным – это тоже типично (что фермент самой медленной реакции метаболического пути является ключевым).
32.7.1. Регуляция гликолиза гормонами.
Гликолиз стимулируется инсулином (за счёт индукции ключевых ферментов) и подавляется глюкокортикостероидами (ГКС; за счёт репрессии ключевых ферментов). См. п.85.
Катехоламины адреналин и норадреналин усиливают гликолиз в мышцах и подавляют в печени.
Благодаря стимуляции инсулином гликолиза (а также стимуляции инсулином транспорта глюкозы во многие клетки) расход глюкозы тканями увеличивается, что приводит
1) к удовлетворению потребностей клеток в глюкозе и
2) к снижению концентрации глюкозы в крови –
это снижение концентрации глюкозы в крови называется гипогликемическим эффектом,
а инсулин, который обусловливает снижение концентрации глюкозы в крови, является гипогликемическим гормоном.
32.7.2. Регуляция гликолиза метаболитами.
Обычно активаторами процесса являются его субстраты, а ингибиторами – продуктами. Механизм – аллостерический (см. п.6).
Активатором гликолиза является АДФ (в качестве субстрата – чтобы превратиться в АТФ), а ингибиторами – продукты гликолиза АТФ, ацетилКоА и цитрат (отдалённо это продукты гликолиза, так как цитрат образуется из ацетилКоА и оксалоацетата, образующихся из пирувата).
Эффекты АДФ противоположны эффектам АТФ, так как АДФ образуется при расщеплении АТФ, и [АДФ] становится тем больше, чем меньше [АТФ].
Ещё один активатор гликолиза – метаболит фруктозо-2,6- бисфосфат, который активирует фосфофруктокиназу.
Сравните регуляцию гликолиза с регуляцией ГНГ – эти два процесса регулируются одними и теми же регуляторами, но действие регуляторов на эти два противоположных процесса прямо противоположно. Это помогает рассказать регуляцию ГНГ, зная регуляцию гликолиза.
Р е г у л я ц и я г о р м о н а м и процессов углеводного обмена.
Усиление процесса
способствует (гипер/гликемии) или не способствует? Г о р м о н ы
Ин ГГ КА ГКС
; [Г] Гипергликемические
Гликолиз (глюкоза в пируват) Нет, ведёт к гипогликемии ; ; в печени см. *
Глюконеогенез (синтез глюкозы) Да, ведёт к гипергликемии ; ; в печени
* В мышцах КА (в т.ч. адреналин) не снижают гликолиз, а усиливают – так как КА должны обеспечить мышцы энергией за счет выработки в мышцах АТФ. Гликолиз – это единственный процесс, который может дать АТФ при дефиците кислорода (дефицит кислорода обязательно возникает в работающих скелетных мышцах, как бы ни старались легкие и сердце обеспечить мышцы кислородом; но чем лучше человек тренирован, тем лучше организм справляется с нагрузками).
Р е г у л я ц и я гликолиза и ГНГ метаболитами. (Аллостерическая).
(Действие регуляторов на гликолиз и на ГНГ противоположно.)
ацетилКоА Цитрат А Т Ф А Д Ф Фруктозо-2,6-бисфосфат
гликолиз Продукт продукт Продукт Субстрат
Ингибиторы гликолиза Активаторы гликолиза
ГНГ: синтез
глюкозы Субстрат продукт
Активаторы ГНГ Ингибиторы ГНГ
Таблица «Гликолиз в разных условиях: в анаэробных и в аэробных». ;
Гликолиз – это «лизис глюкозы», то есть расщепление (катаболизм) глюкозы (ее углеродного скелета) до пирувата,
а далее – в зависимости от условий: в анаэробных условиях пируват превращается в лактат,
а в аэробных поступает в ЦТК и превращается в СО2 и воду (с участием ДЦ).
+ — достоинство процесса, – недостаток.
№ В анаэробных В аэробных
1. Требует ли н а л и ч и я кислорода Нет (это +) Да (это – )
2. Может ли протекать при ишемии, гипоксии (1) Да (это +) нет
3. Нужна ли помощь ЦТК и ДЦ нет да
4. Каким метаболитом заканчивается Лактат (снижает рН, это –) 6 СО2, НОН
5. Приводит ли к образованию лактата Да (это –) Нет (это +)
6. Приводит ли к ацидозу при усилении (см. 5) Да (это –) Нет (это +)
7. Нужно ли у ч а с т и е митохондрий Нет Да
8. Протекает ли в эритроцитах (см. 7) да Нет (т.к. в ЭЦ нет МХ)
9. Сколько АТФ дает (в расчете на 1 молекулу Г) 2 АТФ 32 АТФ (30 в мозге)
10 Достаточен ли для мозга
в качестве источника энергии Нет (поэтому мозг
без О2 гибнет) Да (основной источник
АТФ для мозга)
11 Дает ли метаболиты для ЦТК (ОА и др.) Нет (это –) да (это +)
12
В чем д о с т о и н с т в а
(см. выше) 1) только он дает АТФ в ЭЦ,
2) только он дает АТФ без О2, хоть и мало (всего 2),
3) быстро включается, что ценно при короткой напряженной работе 1) дает много АТФ,
2) дает метаболиты ЦТК
(ОА и ацетилКоА из П,
а затем в ЦТК образуются остальные, в т.ч. для аминокислот и белков)
В чем н е д о с т а т к и
(см. выше) 1) образует лактат, что приводит к ацидозу,
2) мало АТФ (из-за чего жгут нельзя долго держать) не протекает
без кислорода
(при гипоксии)
14 Каким
тканям или клеткам
дает достаточное количество энергии Эритроцитам, скелетным мышцам при короткой работе, а при жгуте – час Всем, в которых протекает
(в эритроцитах не протекает)
О б м е н л а к т а т а . Лактат (Л) – это соль или анион молочной кислоты.
1. Как называется увеличение [Л] в крови Гипер / лактат / емия
2. Чем о п а с н а высокая [ лактата ]
(молочная кислота ; лактат + Н+ ) Накоплением протонов (Н+), т.е. АЦИДОЗОМ, который приводит к
тошноте, усталости и (при очень сильном ацидозе) к коме
3. Какой ПРОЦЕСС образует лактат Анаэробный гликолиз (при дефиците кислорода и в ЭЦ)
4. Какие ТКАНИ являются источником лактата (поступающего в кровь) В покое – эритроциты. При работе скелетных мышц – мышцы.
При патологии – любая ткань при гипоксии и без В1.
5. Какие ПРОЦЕССЫ используют лактат ГНГ (в печени) и окисление лактата (ЛДГ, ПДГ, ЦТК)
6. Какой ОРГАН «берёт» лактат из крови Печень (для превращения лактата в глюкозу в ГНГ)
7.
Причины накопления лактата
(гипер/лактат/емии),
связанные
с увеличением образования лактата
(см. 3 и 4) У здоровых – работа скелетных мышц (усталость, тошнота).
У больных – разные формы Г И П О К С И И , в т.ч:
1. из-за ишемии (жгут, тромб, спазм, падение АД и т.д.),
2. из-за патологии дыхательной системы (эмфизема легких при курении и др.),
3. из-за патологии сердечно-сосудистой системы,
4. из-за анемии (нет Fe, витаминов, белка, есть гемолиз),
5. при шоке, коллапсе, терминальных состояниях.
8. Причины накопления лактата,
связанные с уменьшением его утилизации (использования) 1) патология печени из-за снижения ГНГ
(п/л печени – из-за алкоголизма и др.),
2) дефицит В1, т.к. без него не может работать ПДГ и ЦТК.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом