
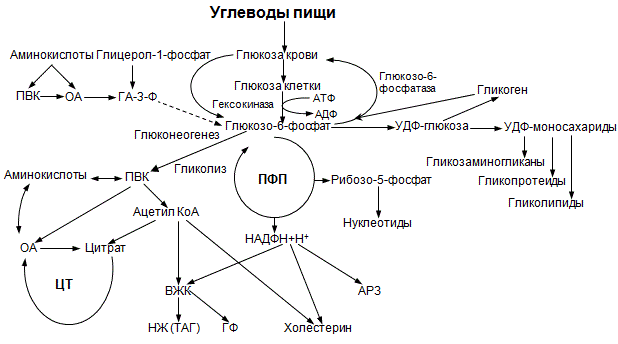
48. Особенности обмена глюкозы в разных органах и клетках: эритроциты, мозг, мышцы, жировая ткань, печень.
Метаболизм глюкозы в эритроцитах. Эритроциты лишены митохондрий, поэтому в качестве энергетического материала они могут использовать только глюкозу. В эритроцитах катаболизм глюкозы обеспечивает сохранение структуры и функции гемоглобина, целостность мембран и образование энергии для работы ионных насосов. Глюкоза поступает в эритроциты путём облегчённой диффузии с помощью ГЛЮТ-2. Около 90% поступающей глюкозы используется в анаэробном гликолизе, а остальные 10% — в пентозофосфатном пути. Конечный продукт анаэробного гликолиза лактат выходит в плазму крови и используется в других клетках, прежде всего гепатоцитах. АТФ, образующийся в анаэробном гликолизе, обеспечивает работу Nа + , К+-АТФ-азы и поддержание самого гликолиза, требующего затраты АТФ в гексокиназной и фосфофруктокиназной реакциях. Важная особенность анаэробного гликолиза в эритроцитах по сравнению с другими клетками — присутствие в них фермента бисфосфоглицератмутазы. Бисфосфоглицератмутаза катализирует образование 2,3-бисфосфоглицерата из 1,3-бисфосфоглицерата. Образующийся только в эритроцитах 2,3-бисфосфоглицерат служит важным аллостерическим регулятором связывания кислорода гемоглобином. Глюкоза в эритроцитах используется и в пентозофосфатном пути, окислительный этап которого обеспечивает образование кофермента NADPH, необходимого для восстановления глу-татиона.
Метаболизм глюкозы в печени.Основная рольпеченив углеводном обмене заключается в обеспечении постоянстваконцентрацииглюкозывкрови. Это достигается регуляцией междусинтезом и распадом гликогена, депонируемого впечени. Впеченисинтез гликогенаи его регуляция в основном аналогичны тем процессам, которые протекают в других органах итканях, в частности вмышечной ткани.Синтез гликогенаизглюкозыобеспечивает в норме временный резервуглеводов, необходимый для поддержанияконцентрацииглюкозывкровив тех случаях, если ее содержание значительно уменьшается (например, у человека это происходит при недостаточном поступленииуглеводовс пищей или в период ночного «голодания»). Необходимо подчеркнуть важную рольферментаглюкокиназы в процессе утилизацииглюкозыпеченью. Глюкокиназа, подобногексокиназе, катализируетфосфорилированиеглюкозыс образованием глюкозо-6-фосфата, при этомактивностьглюкокиназы впеченипочти в 10 раз превышаетактивностьгексокиназы. Важное различие между этими двумяферментамизаключается в том, что глюкокиназа в противоположностьгексокиназеимеет высокое значение КМ дляглюкозыи не ингибируется глюкозо-6-фосфатом. Считают, что основная рольпечени– расщеплениеглюкозы– сводится прежде всего к запасанию метаболитов-предшественников, необходимых длябиосинтеза жирных кислотиглицерина, и в меньшей степени кокислениюее до СО2 и Н2О. Синтезированные впеченитриглицериды в норме выделяются вкровьв составелипопротеинови транспортируются вжировую тканьдля более «постоянного» хранения. Вреакцияхпентозофосфатного пути впечениобразуется НАДФН, используемый для восстановительныхреакцийв процессах синтезажирных кислот,холестеринаи другихстероидов. Кроме того, при этом образуются пентозофосфаты, необходимые для синтезануклеиновых кислот. Наряду с утилизациейглюкозывпеченипроисходит и ее образование. Непосредственным источникомглюкозывпеченислужитгликоген.Распад гликогенавпеченипроисходит в основном фосфоролитическим путем. В регуляции скоростигликогенолизавпеченибольшое значение имеет система циклическихнуклеотидов. Кроме того,глюкозавпечениобразуется также в процессеглюконеогенеза.
Метаболизм глюкозы в мозгу.Основнымсубстратомдыханиямозговойтканиявляетсяглюкоза. В 1 мин 100 гтканимозга потребляют в среднем 5 мгглюкозы. Подсчитано, что более 90% утилизируемойглюкозывтканимозга окисляется до СО2и Н2О при участиицикла трикарбоновых кислот. В физиологических условиях роль пентозофосфатного путиокисленияглюкозыв мозговойтканиневелика, однако этот путьокисленияглюкозыприсущ всемклеткамголовного мозга. Образующаяся в процессепентозофосфатного циклавосстановленная форма НАДФ (НАДФН) используется для синтезажирных кислотистероидов. Междуглюкозойигликогеноммозговойтканиимеется тесная связь, выражающаяся в том, что при недостаточном поступленииглюкозыизкровигликогенголовного мозга является источникомглюкозы, аглюкозапри ее избытке – исходным материалом длясинтеза гликогена.Распад гликогенав мозговойтканипроисходит путем фосфоролиза с участием системыцАМФ. Однако в целом использованиегликогенав мозге по сравнению сглюкозойне играет существенной роли в энергетическом отношении, так как содержаниегликогенав головном мозге невелико. Наряду с аэробнымметаболизмом углеводовмозговаятканьспособна к довольно интенсивному анаэробномугликолизу. Значение этого явления пока недостаточно ясно, ибогликолизкак источник энергии ни в коей мере не может сравниться по эффективности с тканевымдыханиемв головном мозге.
Метаболизм глюкозы в мышцах.При работе умеренной интенсивности мышца может покрывать свои энергетические затраты за счет аэробногометаболизма. Однако при больших нагрузках, когда возможность снабжениякислородомотстает от потребности в нем, мышца вынуждена использовать гликолитический путь снабжения энергией. При интенсивной мышечной работе скорость расщеплениягликогенаилиглюкозыс образованиеммолочной кислотыувеличивается в сотни раз. Соответственно содержаниемолочной кислотывмышечной тканиможет повышаться до 1,0–1,2 г/кг и более. С токомкровизначительное количествомолочной кислотыпоступает впечень, где ресинтезируется вглюкозуигликоген(глюконеогенез) за счет энергии окислительных процессов.
Метаболизм глюкозы в жировой ткани. В адипоцитах для обеспечения реакций синтеза жира распад глюкозы идёт по двум путям: гликолиз, обеспечивающий образование глицерол-3-фосфата и ацетил-КоА, и пентозофосфатный путь, окислительные реакции которого обеспечивают образование NADPH, служащего донором водорода в реакциях синтеза жирных кислот.
Углеводный обмен
Совокупность процессов, протекающих в организме человека, включающих превращение моносахаридов и их производных, гомо-, гетерополисахаридов и различных углеводсодержащих биополимеров носит название углеводного обмена. Его результатом является снабжение организма энергией, осуществление межмолекулярных взаимодействий и процессов передачи биологической информации.
Этапы углеводного обмена
- Пищеварительный этап (переваривание начинается в желудке и заканчивается в тонком кишечнике с участием поджелудочного и кишечного соков, амилазы, лактазы, мальтазы, инвертазы).
- Промежуточный (глюкоза поступает в печень по воротной вене, где происходят гликогенез и неогликогенез, а также гликогенолиз, аналогичные процессы протекают в мышечных тканях).
- Конечный этап (выделение продуктов обмена — воды и углекислого газа из организма).
Причины нарушения углеводного обмена и связанные с ним заболевания
Расстройства метаболизма углеводов делятся:
- на гипогликемии (снижение уровня глюкозы в плазме крови);
- гликогенозы (патологии углеводного обмена врожденного либо наследственного характера, проявляющиеся избыточным накоплением гликогена в клетках организма);
- гексоземии (повышенное содержание гексоз в крови);
- гипергликемии (повышение глюкозы в плазме крови);
- сахарный диабет (нарушения всех звеньев метаболизма углеводов).
Группа нарушений углеводного обмена — гипогликемии, могут быть обусловлены:
- заболеваниями печени (гепатоз, гепатодистрофия, ферментопатии);
- нарушениями пищеварения (проблемы при полостном переваривании углеводов, пристеночном расщеплении и абсорбции);
- болезнями почек (нарушение реабсорбции глюкозы в проксимальных канальцах нефрона);
- эндокринопатиями (глюкокортикоидная недостаточность, дефицит Т3 и Т4, СТГ, катехоламинов, глюкагона);
- углеводным голоданием (из-за нарушений питания);
- длительной и значительной физической работой.
Расстройства углеводного обмена в виде гликогенозов развиваются по причине мутаций генов, которые кодируют синтез ферментов расщепления либо образования гликогена. К таким заболеваниям относят болезнь Гирке, болезнь Помпе, болезнь Андерсена, болезнь Таруи, болезнь Хага и др.
Читайте также: Трикотажная ткань для тельняшек
Нарушения углеводного обмена группы гексоземий включают в себя галактоземию врожденного или наследственного характера, а также фруктоземию с врожденной непереносимостью фруктозы, возникающую по причине недостаточности альдолазы В.
Гипергликемии — расстройства углеводного обмена, которые развиваются по таким причинам:
- эндокринопатии (избыток эффектов гипергликемизирующих факторов и дефицит эффектов инсулина);
- психогенные и неврологические расстройства (психическое возбуждение, стресс-реакции, каузалгии);
- переедание (долгое избыточное употребление сладостей и легкоусвояемых углеводов);
- патологии печени (печеночная недостаточность).
Сахарный диабет — распространенное нарушение углеводного обмена. Подразделяется на инсулинзависимый и инсулиннезависимый. Причинами сахарного диабета считаются:
- дефицит инсулина (генетические дефекты β-клеток островков Лангерганса; вирусы, тропные к β-клеткам; иммунные факторы; эндогенные токсические вещества; воспалительные процессы; физические и химические факторы);
- недостаточность эффектов инсулина (нейро- или психогенных факторы; контринсулярных факторы; дефекты инсулиновых рецепторов и пострецепторные нарушения в клетках-мишенях).
Лечение и профилактика нарушений углеводного обмена
Терапия направлена на нормализацию метаболизма углеводов в организме. Лечение разрабатывается индивидуально исходя из типа и выраженности патологии углеводного обмена.
Профилактика нарушений метаболизма заключается в правильном сбалансированном питании, своевременном лечении возникающих заболеваний печени, почек, эндокринных желез.
Узнать индивидуальные риски по развитию болезней, связанных с углеводным обменом, позволяет генетическое тестирование. В медико-генетическом центре «Геномед» можно пройти исследование «Наследственные нарушения обмена веществ».
Особенности обмена глюкозы в разных органах и клетках: эритроциты, мозг, мышцы, жировая ткань, печень.
Метаболизм глюкозы в эритроцитах. Эритроциты лишены митохондрий, поэтому в качестве энергетического материала они могут использовать только глюкозу. В эритроцитах катаболизм глюкозы обеспечивает сохранение структуры и функции гемоглобина, целостность мембран и образование энергии для работы ионных насосов. Глюкоза поступает в эритроциты путём облегчённой диффузии с помощью ГЛЮТ-2. Около 90% поступающей глюкозы используется в анаэробном гликолизе, а остальные 10% — в пентозофосфатном пути. Конечный продукт анаэробного гликолиза лактат выходит в плазму крови и используется в других клетках, прежде всего гепатоцитах. АТФ, образующийся в анаэробном гликолизе, обеспечивает работу Nа+, К+-АТФ-азы и поддержание самого гликолиза, требующего затраты АТФ в гексокиназной и фосфофруктокиназной реакциях. Важная особенность анаэробного гликолиза в эритроцитах по сравнению с другими клетками — присутствие в них фермента бисфосфоглицератмутазы. Бисфосфоглицератмутаза катализирует образование 2,3-бисфосфоглицерата из 1,3-бисфосфоглицерата. Образующийся только в эритроцитах 2,3-бисфосфоглицерат служит важным аллостерическим регулятором связывания кислорода гемоглобином. Глюкоза в эритроцитах используется и в пентозофосфатном пути, окислительный этап которого обеспечивает образование кофермента NADPH, необходимого для восстановления глу-татиона.
Метаболизм глюкозы в печени. Основная роль печени в углеводном обмене заключается в обеспечении постоянства концентрации глюкозы в крови. Это достигается регуляцией между синтезом и распадом гликогена, депонируемого в печени. В печени синтез гликогена и его регуляция в основном аналогичны тем процессам, которые протекают в других органах и тканях, в частности в мышечной ткани. Синтез гликогена из глюкозы обеспечивает в норме временный резерв углеводов, необходимый для поддержания концентрации глюкозы в крови в тех случаях, если ее содержание значительно уменьшается (например, у человека это происходит при недостаточном поступлении углеводов с пищей или в период ночного «голодания»). Необходимо подчеркнуть важную роль фермента глюкокиназы в процессе утилизации глюкозы печенью. Глюкокиназа, подобногексокиназе, катализирует фосфорилирование глюкозы с образованием глюкозо-6-фосфата, при этом активность глюкокиназы в печенипочти в 10 раз превышает активность гексокиназы. Важное различие между этими двумя ферментами заключается в том, что глюкокиназа в противоположность гексокиназе имеет высокое значение КМ для глюкозы и не ингибируется глюкозо-6-фосфатом. Считают, что основная роль печени – расщепление глюкозы – сводится прежде всего к запасанию метаболитов-предшественников, необходимых для биосинтеза жирных кислот и глицерина, и в меньшей степени к окислению ее до СО2 и Н2О. Синтезированные впечени триглицериды в норме выделяются в кровь в составе липопротеинов и транспортируются в жировую ткань для более «постоянного» хранения. В реакциях пентозофосфатного пути в печени образуется НАДФН, используемый для восстановительных реакций в процессах синтезажирных кислот, холестерина и других стероидов. Кроме того, при этом образуются пентозофосфаты, необходимые для синтезануклеиновых кислот. Наряду с утилизацией глюкозы в печени происходит и ее образование. Непосредственным источником глюкозы в печени служитгликоген. Распад гликогена в печени происходит в основном фосфоролитическим путем. В регуляции скорости гликогенолиза в печенибольшое значение имеет система циклических нуклеотидов. Кроме того, глюкоза в печени образуется также в процессе глюконеогенеза.
Метаболизм глюкозы в мозгу. Основным субстратом дыхания мозговой ткани является глюкоза. В 1 мин 100 г ткани мозга потребляют в среднем 5 мг глюкозы. Подсчитано, что более 90% утилизируемой глюкозы в ткани мозга окисляется до СО2 и Н2О при участии цикла трикарбоновых кислот. В физиологических условиях роль пентозофосфатного пути окисления глюкозы в мозговой ткани невелика, однако этот путь окисленияглюкозы присущ всем клеткам головного мозга. Образующаяся в процессе пентозофосфатного цикла восстановленная форма НАДФ (НАДФН) используется для синтеза жирных кислот и стероидов. Между глюкозой и гликогеном мозговой ткани имеется тесная связь, выражающаяся в том, что при недостаточном поступлении глюкозыиз крови гликоген головного мозга является источником глюкозы, а глюкоза при ее избытке – исходным материалом для синтеза гликогена. Распад гликогена в мозговой ткани происходит путем фосфоролиза с участием системы цАМФ. Однако в целом использование гликогена в мозге по сравнению с глюкозой не играет существенной роли в энергетическом отношении, так как содержание гликогена в головном мозге невелико. Наряду с аэробным метаболизмом углеводов мозговая ткань способна к довольно интенсивному анаэробному гликолизу. Значение этого явления пока недостаточно ясно, ибо гликолиз как источник энергии ни в коей мере не может сравниться по эффективности с тканевым дыханием в головном мозге.
Читайте также: Как отстирать жирное пятно с синтетической ткани
Метаболизм глюкозы в мышцах. При работе умеренной интенсивности мышца может покрывать свои энергетические затраты за счет аэробного метаболизма. Однако при больших нагрузках, когда возможность снабжения кислородом отстает от потребности в нем, мышца вынуждена использовать гликолитический путь снабжения энергией. При интенсивной мышечной работе скорость расщепления гликогена или глюкозы с образованием молочной кислоты увеличивается в сотни раз. Соответственно содержание молочной кислоты в мышечной ткани может повышаться до 1,0–1,2 г/кг и более. С током крови значительное количество молочной кислоты поступает в печень, где ресинтезируется в глюкозу и гликоген (глюконеогенез) за счет энергии окислительных процессов.
Метаболизм глюкозы в жировой ткани. В адипоцитах для обеспечения реакций синтеза жира распад глюкозы идёт по двум путям: гликолиз, обеспечивающий образование глицерол-3-фосфата и ацетил-КоА, и пентозофосфатный путь, окислительные реакции которого обеспечивают образование NADPH, служащего донором водорода в реакциях синтеза жирных кислот.
49. Протеогликаны— высокомолекулярные соединения, состоящие из белка (5-10%) и гликозаминогликанов (90-95%). Они образуют основное вещество межклеточного матрикса соединительной ткани и могут составлять до 30% сухой массы ткани.
Cтроение и виды протеогликанов. В межклеточном матриксе присутствуют разные протеогликаны. Среди них есть очень крупные — например агрекан и версикан. Кроме них, в межклеточном матриксе имеется целый набор так называемых малых протеогликанов, которые широко распространены в разных видах соединительной ткани и выполняют там самые разнообразные функции. Основной протеогликан хрящевого матрикса называется агрекан, он составляет 10% по весу исходной ткани и 25% сухого веса хрящевого матрикса. Это очень большая молекула, в которой к одной полипептидной цепи присоединены до 100 цепей хондроитинсульфатов и около 30 цепей кератансульфатов. По форме молекула агрекана напоминает бутылочный «ёршик». В хрящевой ткани молекулы агрекана собираются в агрегаты с гаалуроновой кислотой и небольшим связывающим белком. Оба компонента присоединяются к агрекану нековалент-ными связями в области домена G1. Домен G1 взаимодействует примерно с пятью дисахаридными единицами гиалуроновой кислоты, далее этот комплекс стабилизируется связывающим белком; домен G1 и связывающий белок вместе занимают 25 дисахаридных единиц гиалуроновой кислоты. Конечный агрегат с молекулярной массой более 200 × 106 Д состоит из одной молекулы гиалуроновой кислоты и 100 молекул агрекана (и такого же количества связывающего белка). Координация сборки этих агрегатов является центральной функцией хондроцитов. Агрекан и связывающий белок продуцируются этими клетками в необходимых количествах. Эти компоненты могут взаимодействовать друг с другом внутри клетки, но процесс агрегации полностью завершается в межклеточном матриксе. Показано, что гиалуроновая кислота образуется на поверхности хондроцитов специфической синтетазой и «выталкивается» в межклеточное пространство, чтобы связаться с агреканом и связывающим белком. Созревание функционально активного тройного комплекса составляет около 24 ч.
Малые протеогликаны. Малые протеогликаны — протеогликаны с низкой молекулярной массой. Они содержатся в хрящах, сухожилиях, связках, менисках, коже и других видах соединительной ткани. Эти протеогликаны имеют небольшой коровый белок, к которому присоединены одна или две цепи гликозаминогликанов. Наиболее изучены декорин, бигликан, фибромодулин, люмикан, перлекан. Коровые белки бигликана и декорина похожи по размерам и структуре (молекулярная масса 36 000 и 38 000 Д, соответственно). Они имеют несколько тандемных повторов, богатых лейцином, которые образуют α-спирали или β-структуры. На N- и С-концах этих белков имеются домены, содержащие S-S-связи. Ко’ровые белки значительно различаются по первичной структуре в N-концевых областях, что определяет различия в присоединении гликозаминогликанов. Бигликан содержит серии в положениях 5 и 11, что обеспечивает присоединение двух полисахаридных цепей. Декорин содержит один серии в положении 4, поэтому к нему присоединяется одна полисахаридная цепь. У этих протеогликанов полисахаридные цепи представлены дерматансульфатом с молекулярной массой
30 000 Д.Ко́ровый белок фибромодулина (молекулярная масса
40 000 Д) тоже имеет области тандемных повторов, богатые лейцином, но его N-концевая область отличается тем, что не содержит серина, а имеет несколько сульфатированных остатков тирозина, поэтому одна или две цепи кератансульфата присоединяются к ко’ровому белку фибромодулина не в N-концевой, а в области, богатой лейцином, через NH2-группу аспарагина. Малые протеогликаны являются мультифункциональными макромолекулами. Они могут связываться с другими компонентами соединительной ткани и оказывать влияние на их строение и функции. Например, декорин и фибромодулин присоединяются к фибриллам коллагена II типа и ограничивают их диаметр (т.е. препятствуют образованию толстых фибрилл). Декорин и бигликан, присоединяясь к фибронектину, подавляют клеточную адгезию, а присоединяясь к фактору роста опухолей (3, снижают его митогенную активность. Кроме этого, имеется большое количество данных о том, что малые протеогликаны играют важную регуляторную роль в процессах развития и восстановления соединительной ткани.
- они являются структурными компонентами межклеточного матрикса;
- протеогликаны и гликозаминогликаны специфически взаимодействуют с коллагеном, эластином, фибронектином, ламинином и другими белками межклеточного матрикса;
- все протеогликаны и гликозаминогликаны, являясь полианионами, могут присоединять, кроме воды, большие количества катионов (Na+, K+, Са 2+ ) и таким образом участвовать в формировании тургора различных тканей;
- протеогликаны и гликозаминогликаны играют роль молекулярного сита в межклеточном матриксе, они препятствуют распространению патогенных микроорганизмов;
- гиалуроновая кислота и протеогликаны выполняют рессорную функцию в суставных хрящах;
- гепарансульфатсодержащие протеогликаны способствуют созданию фильтрационного барьера в почках;
- кератансульфаты и дерматансульфаты обеспечивают прозрачность роговицы;
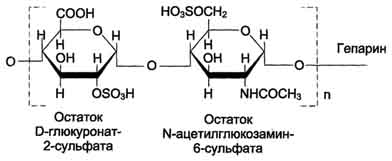
- гепарин — антикоагулянт;
- гепарансульфаты — компоненты плазматических мембран клеток, где они могут функционировать как рецепторы и участвовать в клеточной адгезии и межклеточных взаимодействиях. Они также выступают компонентами синаптических и других пузырьков.
50. Гликозаминогликаны представляют собой длинные неразветвлённые цепи гетерополиса-харидов. Они построены из повторяющихся дисахаридных единиц. Одним мономером этого дисахарида является гексуроновая кислота (D-глюкуроновая кислота или L-идуроновая), вторым мономером — производное аминосахара (глюкоз- или галактозамина). NH2-rpynna аминосахаров обычно ацетилирована, что приводит к исчезновению присущего им положительного заряда. Кроме гиалуроновой кислоты, все гликозаминогликаны содержат сульфатные группы в виде О-эфиров или N-сульфата.
Читайте также: Светло серая костюмная ткань

и других пузырьков
Хондроитинсульфаты— самые распространённые гликозаминогликаны в организме человека; они содержатся в хряще, коже, сухожилиях, связках, артериях, роговице глаза. Хондроитинсульфаты являются важным составным компонентом агрекана — основного протеогликана хрящевого матрикса. В организме человека встречаются 2 вида хондроитинсульфатов: хондроитин-4-сульфат и хондроитин-6-сульфат. Они построены одинаковым образом, отличие касается только положения сульфатной группы в молекуле N-ацетилгалактозамина.

Кератансульфаты— наиболее гетерогенные гликозаминогликаны; отличаются друг от друга по суммарному содержанию углеводов и распределению в разных тканях. Кератансульфат I находится в роговице глаза и содержит кроме повторяющейся дисахаридной единицы L-фукозу, D-маннозу и сиаловую кислоту. Кератансульфат II был обнаружен в хрящевой ткани, костях, межпозвоночных дисках

Дерматансульфатшироко распространён в тканях животных, особенно он характерен для кожи, кровеносных сосудов, сердечных клапанов.

Гепарин— важный компонент противосвёртывающей системы крови (его применяют как антикоагулянт при лечении тромбозов).
51. . Функции и особенности обмена гликозаминогликанов
• они являются структурными компонентами межклеточного матрикса;
• протеогликаны и гликозаминогликаны специфически взаимодействуют с коллагеном, эластином, фибронектином, ламинином и другими белками межклеточного матрикса;
• все протеогликаны и гликозаминогликаны, являясь полианионами, могут присоединять, кроме воды, большие количества катионов (Na+, K+, Са2+) и таким образом участвовать в формировании тургора различных тканей;
• протеогликаны и гликозаминогликаны играют роль молекулярного сита в межклеточном матриксе, они препятствуют распространению патогенных микроорганизмов;
• гиалуроновая кислота и протеогликаны выполняют рессорную функцию в суставных хрящах;
• гепарансульфатсодержащие протеогликаны способствуют созданию фильтрационного барьера в почках;
• кератансульфаты и дерматансульфаты обеспечивают прозрачность роговицы;
• гепарансульфаты — компоненты плазматических мембран клеток, где они могут функционировать как рецепторы и участвовать в клеточной адгезии и межклеточных взаимодействиях. Они также выступают компонентами синаптических
52. Олигосахаридный компонент гликопротеинов и гликолипидов. Строение, биологическая роль.
Гликопротеины – сложные белки, содержащие, помимо простого белка или пептида, группу гетероолигосахаридов. В настоящее время их принято называть гликоконъюгатами. В состав гликоконъюгата входит углеводный компонент (гликановая фракция), ковалентно связанный с неуглеводной частью (агликановая фракция), представленной белком, пептидом, аминокислотой или липидом. Химический состав гликопротеинов более или менее установлен, структура определена только у ряда из них. К полипептидуприсоединяются гетероолигосахаридные цепи, содержащие от 2 до 10, реже 15 мономерных остатков гексоз (галактоза и манноза, режеглюкоза), пентоз (ксилоза, арабиноза) и конечный углевод, чаще всего представленный N-ацетилга-лактозамином, L-фукозой или сиаловой кислотой; в отличие от про-теогликанов гликопротеины не содержат уроновых кислот и серной кислоты. Типы связей между углеводными компонентами и белками определены только у ряда гликопротеинов, аминокислотный состав и структура которых известны (иммуноглобулины, гормоны); они включают О-гликозидные связи (с ОН-группами серина, треонина и оксилизина), N-гликозидные связи (с амидными группами аспарагина, реже глутамина или ω-NH2-группами лизина и аргинина) и эфирные гликозидные связи со свободными СООН-группами глутаминовой и аспарагиновой кислот. Углеводные компоненты соединены ковалентно с азотом аспарагина молекулы белка.
Гликолипиды —сложные липиды, образующиеся в результате соединения липидов с углеводами. У гликолипидов имеются полярные «головы» (углевод) и неполярные «хвосты» (остатки жирных кислот). Благодаря данному свойству, вместе с фосфолипидами, гликолипиды входят в состав клеточных мембран. Гликолипиды широко представлены в тканях, особенно в нервной ткани, в частности в ткани мозга. Они локализованы преимущественно на наружной поверхности плазматической мембраны, где их углеводые компоненты входят в число других углеводов клеточной поверхности.
53. Сиаловые кислоты. Основные представители, содержание в крови и тканях в норме и при патологии.
Сиа́ловые кисло́ты — ациальные производные нейраминовой кислоты, присутствуют во всех тканях и жидкостях организма животных и человека и у некоторых микроорганизмов. У человека в норме наибольшее количество Сиаловые кислоты обнаруживается в слюнных железах, в секретах различных слизистых оболочек, а также в сыворотке крови, где их содержание резко повышается при ряде заболеваний. Сиаловые кислоты являются полифункциональными соединениями с сильными кислотными свойствами. Как правило, в свободном виде в норме они не встречаются, а входят в состав различных углеводсодержащих веществ, таких как гликопротеины, гликолипиды (ганглиозиды), олигосахариды. Занимая в молекулах этих веществ концевое положение, сиаловые кислоты оказывают значительное влияние на их физико-химические свойства и биологическую активность. Определяя отрицательный заряд молекул гликопротеинов, сиаловые кислоты обусловливают вытянутую форму их молекул и как следствие высокую вязкость содержащих эти гликопротеины секретов слизистых оболочек дыхательного, кишечного и полового трактов. Это обеспечивает защиту слизистых оболочек от механических и химических повреждений. Наличие сиаловых кислот в составе белков крови (церулоплазмина, кислого α1-гликопротеина и др.) и некоторых гормонов (хорионического гонадотропина, фолликулостимулирующего и лютеинизирующего гормонов) определяет длительность циркуляции этих соединений в кровотоке. После отщепления сиаловой кислоты, когда концевым сахаром в молекулах гликопротеинов становится галактоза, эти белки поглощаются клетками печени. Именно этим объясняется потеря гормонами биологической активности. Длительность циркуляции в кровотоке некоторых клеток крови (эритроцитов, лимфоцитов) также зависит от наличия или отсутствия сиаловых кислот на их поверхности. Процесс старения эритроцитов связан с уменьшением количества сиаловых кислот в их оболочке. Находясь в составе углеводной части гликопротеинов, сиаловые кислоты маскируют остатки сахаров, являющихся антигенными детерминантами и таким образом, играют важную роль в иммунных реакциях, снижая иммуногенные свойства нормальных и опухолевых клеток. Установлено, что сиаловые кислоты являются компонентом клеточных рецепторов, специфичных для вирусов гриппа.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом
- Правообладателям
- Политика конфиденциальности