
Физиологические особенности мембран возбудимых тканей. Роль
Общая физиология возбудимых тканей
Раздражимость и возбудимость. Виды возбудимых тканей и их свойства. Общие и специфические признаки возбуждения. Законы возбуждения (силы, времени и градиента). Классификация раздражителей.
Раздражимость – способность реагировать изменением обмена веществ на действие раз-ля
Возбудимость – ответ на раз-е возбуждением
Возбудимые ткани: нервные мышечные, некоторые секреторные
Возбуждение — -ответ на раздражение
Специализированные признаки возбуждения: проведение возбуждения нервной тканью, сокращение мышцы, секреция железы
Общие признаки возбуждения: Генерация ПД, метаболитические изменения.
Раздражители: 1) Естественные (нервные импульсы)
2)Искуственные (физ., хим.,температура ток..)
Адекватные – при минимальных энергозатратах вызывают возбуждение в естественных условиях
Неадекватные – вызывают возбуждение при достаточной силе и длительности воздействия
1) Закон силы: Чем больше сила раздражителя, тем сильнее ответная реакция. Однако выраженность ответной реакции растет лишь до определенного максимума. Закону силы подчиняется целостная скелетная, гладкая мышца, так как они состоят из многочисленных мышечных клеток, имеющих различную возбудимость.
2) .Закон силы-длительности.(времени) Между силой и длительностью действия раздражителя имеется определенная взаимосвязь. Чем сильнее раздражитель, тем меньшее время требуется для возникновения ответной реакции. Зависимость между пороговой силой и необходимой длительностью раздражения отражается кривой силы-длительности. По этой кривой можно определить ряд параметров возбудимости.
а) Порог раздражения – это минимальная сила раздражителя, при которой возникает возбуждение.
б) Реобаза – это минимальная сила раздражителя, вызывающая возбуждение при его действии в течение неограниченно долгого времени. На практике порог и реобаза имеют одинаковый смысл. Чем ниже порог раздражения или меньше реобаза, тем выше возбудимость ткани.
в) Полезное время – это минимальное время действия раздражителя силой в одну реобазу, за которое возникает возбуждение.
г) Хронаксия – это минимальное время действия раздражителя силой в две реобазы, необходимое для возникновения возбуждения. Этот параметр предложил рассчитывать Л. Лапик, для более точного определения показателя времени на кривой силы-длительности. Чем короче полезное время или хронаксия, тем выше возбудимость и наоборот.
В клинической практике реобазу и хронаксию определяют с помощью метода хронаксиметрии для исследования возбудимости нервных стволов.
Закон градиента или аккомодации. Реакция ткани на раздражение зависит от его градиента, Т.е. чем быстрее нарастает сила раздражителя во времени тем быстрее возникает ответная реакция. При низкой скорости нарастания силы раздражителя растет порог раздражения. Поэтому если сила раздражителя возрастает очень медленно, возбуждения не будет. Это явление называется аккомодацией.
Физиологические особенности мембран возбудимых тканей. Роль
мембранных белков. Виды и значение ионных каналов и насосов в функционировании возбудимых клеток. Механизмы деятельности Na + -K + -насоса, Са 2+ -насоса и Na + -Ca 2+ -насоса. Ионные каналы и насосы как мишени действия лекарств.
Биологические мембраны — функционально активные структуры клеток, ограничивающие цитоплазму и большинство внутриклеточных структур; образуют единую внутриклеточную систему канальцев, складок и замкнутых полостей. Структурная основа мембраны – двойной слой фосфолипидов, в который встроены мембранные белки.
Молекулы липидов амфотерны. Своими гидрофильными частями они обращены в сторону водной среды (межклеточная жидкость и цитоплазма), гидрофобные части молекул направлены внутрь липидного бислоя.
Белковые молекулы выполняют роль каналов рецепторов, насосов, ферментов.
Канал – это белковая макромолекула, образующая пору через двухслойную липидную мембрану.
Натриевый канал – тетродотоксин. Калиевый канал – тетраэтиламмоний.
Пора — молекулярное динамическое образование. Образована транспортным ферментом, который способен в 200 раз ускорить диффузию.
Воротный механизм – на внутренней стороне мембране, это белок, способный к конформации (изменение пространственной конфигурации молекул)
Ионные каналы мембраны – Белковые трубочки разного диаметра заполненные жидкостью проходящие мембрану насквозь
Сенсор напряжения – белковая молекула в самой мембране, способна реагировать на изменение мембранного потенциала.
Селективный фильтр – определяет однонаправленное движение ионов через пору и ее избирательную проницаемость.
Пассивный перенос веществ через мембраны проходит без затрат энергии при наличии градиентов (разницы концентраций веществ, разности электрохимического градиента, при наличие градиента давления и осмотического градиента). При этом пассивный транспорт осуществляется с помощью:
— фильтрация. Осуществляется при наличии разности гидростатического давления.
— осмос. При осмосе происходит движение растворителя. То есть вода из чистого раствора будет переходить в раствор с большей концентрацией.
Во всех этих случаях не происходит затраты энергии. Вещества идут через поры, которые имеются в мембране.
В мембране существуют поры с медленной проводимостью, но таких пор в мембране не много. Большинство каналов в мембране имеет в своем строении еще воротный механизм, который перекрывает канал. Эти каналы могут управляться двумя способами: реагировать на изменение заряда (электровозбудимые или потенциалозависимые каналы). В другом случае ворота в канале открываются, когда присоединяется химическое вещество (хемовозбудимые или лигандозависимые).
Активный перенос веществ через мембрану связан с переносом веществ против градиента.
Для активного транспорта используются интегральные белки, которые обладают ферментативными функциями. В качестве энергии используется АТФ. Интегральные белки имеют специальные механизмы (белок), которые активизируется либо при повышении концентрации вещества снаружи клетки, или при понижении внутри.
Наиболее важный процесс активного транспорта — это работа Na/K-насоса, существующего практически во всех клетках; насос выкачивает ионы натрия из клетки, одновременно накачивая ионы калия внутрь клетки. Таким образом обеспечивается низкая внутриклеточная концентрация ионов натрия и высокая-калия (табл. 1.1). Градиент концентрации ионов натрия на мембране имеет специфические функции, связанные с передачей информации в виде электрических импульсов, а также с поддержанием других активных транспортных механизмов и регулирования объема клетки (см. ниже). Поэтому неудивительно, что более 1/3 энергии, потребляемой клеткой, расходуется на Na/К-насос, а в некоторых наиболее активных клетках на его работу расходуется до 70% энерги
Читайте также: Как завязать юбку из куска ткани
насос—кальциевый; это насос откачивает ионы кальция (Са2+) из клетки и участвует в поддержании их внутриклеточной концентрации на крайне низком уровне (табл. 1.1). Кальциевый насос присутствует с очень высокой плотностью в саркоплазматическом рети-кулуме мышечных клеток, которые накапливают ионы кальция в результате расщепления молекул АТФ.
Ионные каналы — порообразующие белки (одиночные либо целые комплексы), поддерживающие разницу потенциалов, которая существует между внешней и внутренней сторонами клеточной мембраны всех живых клеток. Относятся к транспортным белкам. С их помощью ионы перемещаются согласно их электрохимическим градиентам сквозь мембрану. Такие комплексы представляют собой набор идентичных или гомологичных протеинов, плотно упакованных в липидном бислое мембраны вокруг водной поры. Каналы расположены в клеточной и некоторых внутренних мембранах клетки.
Через ионные каналы проходят ионы Na+ (натрия), K+ (калия), Cl- (хлора) и Ca++ (кальция). Из-за открывания и закрывания ионных каналов меняется концентрация ионов по разные стороны мембраны и происходит сдвиг мембранного потенциала.
Строение и физиологические функции мембраны клеток возбудимых тканей
Клетки возбудимых тканей отделены от интерстициальной среды клеточной мембраной. Мембрана нервной и мышечной клетки представляет собой эластичную структуру, толщиной 6—8 нм, состоящую из липидов и белков. Липиды мембраны возбудимых клеток представлены различными фосфолипидами: фосфатидилхолин, фосфатидилэтаноламин, фосфатидил- серин, сфингомиелин и фосфатидилинозит. Фосфолипиды образуют двойной слой, который состоит из фосфолипидных молекул. Полярные головки этих молекул обращены наружу. К каждой полярной головке прикреплены две углеводородные цепочки, которые погружены в толщу мембраны (рис. 2.1).Обращенные наружу полярные головки фосфолипидных молекул клеточных мембран придают им гидрофильные свойства, а удерживаю-
Ионный канал Ионный насос ‘ Наружная
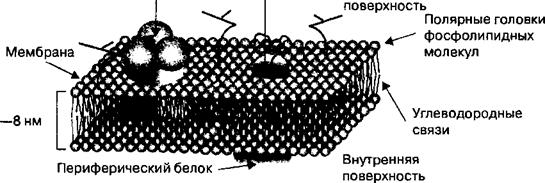
Рис. 2.1.Схематическое изображение мембраны клетки возбудимых тканей.
Мембрана представляет собой двойной липидный слой, в который погружены интегральные белки, функционирующие как ионные насосы и ионные каналы. Ионные насосы за счет энергии АТФ перекачивают ионы К + , Na + , Са 2+ против их концентрационного градиента. Наличие градиентов концентрации указанных ионов на мембране возбудимых клеток является основой возникновения на мембране клеток электрического потенциала и возбудимости нервной и мышечной ткани. Ионные каналы мембраны возбудимой клетки в зависимости от их зактытого или открытого состояния обусловливают возникновение возбуждения в нервной и мышечной ткани и его распространение по поверхности мембран клеток. Периферические белки расположены, как правило, на внутренней поверхности мембраны и являются энзимами, участвующими в регуляции метаболических процессов в возбудимых клетках.
щие их довольно жестко связи углеводородных цепей — гидрофобные. Белки мембран возбудимых клеток подразделяются на два типа: интегральные и периферические. Молекулы интегральных белков погружены в толщу мембраны на большую или меньшую глубину. Этот тип белков обеспечивает взаимодействие (интеграцию) между клетками возбудимых тканей. Интегральные белки выполняют функцию специальных ионных насосов в мембране клеток. Например, с помощью ионных насосов различные ионы (Na\ К + , Са 2+ и др.) перемещаются через мембрану между интерстициальной и внутриклеточной средами, что лежит в основе формирования электрических явлений в возбудимых клетках. Периферические белки расположены, как правило, с внутренней стороны мембраны. Они являются катализаторами протекающих в клетке химических реакций, т. е. являются энзимами. Периферические белки формируют цитоскелет клеток, который придает мембране клеток прочность и одновременно гибкоэластические свойства.
Мембрана клеток возбудимых тканей имеет функции избирательной проницаемости для ионов и веществ, межклеточного взаимодействия, а также экзо- и эндоцитоза, благодаря которым в нервной и мышечной ткани осуществляются физиологические процессы возбуждения, проведения возбуждения и сокращения.
2.1.1. Транспорт веществ через клеточную мембрану
Клеточная мембрана является разделительным барьером между цитоплазмой и внеклеточной средой. При этом важнейшая функция подобного барьера связана с транспортом воды, различных ионов (Na + , К + , СГ, Са 2 * и др.) и питательных веществ (например, глюкозы) между этими средами с целью обеспечения жизнедеятельности клеток.
2.1.1.1. Движение воды через мембрану клеток
Вода довольно свободно проникает через мембрану большинства клеток, так как внутри клеточной мембраны имеются свободные пространства, обусловленные временными дефектами или кинками в СН2 цепях фосфолипидов. Кинки и связанные с ними пространства свободно диффундируют в пределах мембраны. Вода, заключенная в свободно диффундирующие пространства, способна проникать с их помощью через биологические мембраны. Благодаря кинкам и свободным пространствам внутри мембран их «водная» проницаемость примерно в три раза выше, чем в мембранах, организованных только из кристаллической липидной фазы. Однако движение воды через мембрану внутрь клетки и из клетки наружу регулируется осмотическим давлением растворов по обе ее стороны.
Вода диффундирует через мембрану из области меньшей в область большей концентрации растворенного вещества. Этот процесс называется осмосом. Наименьшее гидростатическое давление, которое необходимо приложить к раствору для того, чтобы предотвратить перетекание растворителя через полупроницаемую мембрану в раствор с большей концентрацией вещества называется осмотическим давлением. Концентрация осмотически активных молекул в единице массы воды называется осмолярностью или осмотическим коэффициентом. Осмотическое давление обусловливает поступление воды в клетку, формирует ее объем и упругость (тургор) ткани.
Читайте также: Образцы тканей по цветам
Тоничность раствора представляет собой осмотическое давление, производимое раствором через проницаемую мембрану, отделяющую его от плазмы крови. Терминами изо-, гипо- и гипертонический характеризуют растворы, осмолярность которых соответственно равна, меньше или выше, чем таковая в плазме крови.
Процесс свободного движения молекул вещества в пределах раствора называется диффузией. Большие молекулы движутся в растворе медленнее, чем маленькие, а движение молекул вещества в воде происходит быстрее, чем в среде с более высокой вязкостью. Молекулы биологически важных веществ (медиаторы, гормоны), О2 и СО2 при температуре окружающей среды 30 °C преодолевают расстояние 1 мкм в течение 1 мс, 100 мкм —в течение 10 с, а расстояние 1 см —в течение 24 ч. В клетках возбудимых тканей диффузия молекул в цитоплазме лимитирована их размером. В интерстициальной среде расстояние диффузии молекул веществ ограничено дистанцией между клетками либо между клетками и капиллярами. Так, в среднем расстояние между капиллярами и клетками тканей у млекопитающих животных составляет 30 мкм. Поэтому небольшие по размерам молекулы О2 и СО2 преодолевают это расстояние в течение двух-трех десятков миллисекунд, что обеспечивает обмен газами между кровью и тканями организма.
Диффузия молекул вещества через проницаемую мембрану осуществляется по концентрационному градиенту вещества и описывается первым законом Фика. Подобная диффузия носит название простой и ее величина тем больше, чем выше температура раствора. Первый закон диффузии Фика, который отражает скорость диффузии вещества через мембрану в зависимости от разницы концентраций вещества по обе ее стороны, выражается следующей формулой:
где D — коэффициент диффузии, А —площадь мембраны (см 2 ), х —толщина мембраны (см), С — концентрация вещества по обе стороны мембраны (i — внутри, е — снаружи). С помощью простой диффузии большинство субстанций не проникают через мембрану клетки. Поэтому, наряду с простой диффузией, имеет место движение веществ через мембрану с помощью облегченной диффузии. Облегченная диффузия происходит при участии специфических белковых молекул-переносчиков. Скорость облегченной диффузии лимитируется количеством транспортных белковых молекул и их подвижностью в пределах мембраны. Этот вид мембранного транспорта имеет более высокую скорость в сравнении с простой диффузией. Перенос веществ через мембрану с помощью облегченной диффузии обусловлен строением переносимых молекул, которые связываются с белком-переносчиком. Ингибиторы способны подавлять этот вид транспорта. Для облегченной диффузии характерна специфичность белков-переносчиков к типу молекулы переносимого вещества и количеством транспортных белков в мембране клетки. Например, перенос глюкозы или аминокислот осуществляется строго специфичными белками-переносчиками. Вещества, которые транспортируются через мембрану одним и тем же белком-переносчиком, конкурируют за переносчик (явление конкуренции). Если все белки-переносчики заняты транспортируемыми молекулами или ионами, то увеличение градиента концентрации вещества по обе стороны мембраны не приведет к росту диффузии — явление насыщения.
2.1.1.4. Первично-активный транспорт
Перенос ионов через клеточную мембрану может происходить против их градиента концентрации за счет энергии макроэргических соединений (АТФ). Транспорт ионов через мембрану возбудимых клеток против концентрационного градиента, обусловленный функцией транспортных АТФаз называется первично активным. Первично активный транспорт характерен для переноса через мембрану возбудимых клеток ионов Na + , К + или Са 2+ . Транспортные АТФазы —- это интегральный тип белков клеточной мембраны, поэтому ионы могут активно транспортироваться, например, из внеклеточной среды в цитоплазму (ионы К + ) или наоборот (ионы Na + ).
В клетках возбудимых тканей первично активный транспорт осуществляется с помощью трех типов АТФаз: натрий-калиевой, кальциевой и протонной помпы.
Транспорт ионов Na + и К + через мембрану. В мембранах всех клеток организма человека и животных локализована Иа + /К + -АТФаза, или натрий-калиевый насос(рис. 2.2).Функция этого насоса заключается в поддержании градиента концентрации ионов Na + и К? между цитозолем клетки и внеклеточной средой. В возбудимых клетках создание и поддержание подобного градиента является основным условием возникновения потенциала покоя на мембране клеток, а также последующей генерации и распространения потенциала действия по мембране нервного волокна и мышечной клетки.
 |
| Рис. 2.2.Натрий-калиевый насос (Na + / К + -АТФаза) в мембране возбудимой клетки. |
 |
| Рис. 2.3.Схематическое изображение цикла работы электрогенного натрий-калиевого насоса. |
Конформация Ej насоса обращена в цитозоль и имеет на поверхности два отрицательно заряженных места, с которыми у внутриклеточных ионов Na + и К + имеется различное сродство. Третье электрически нейтральное место связывания ионов натрия находится внутри молекулы насоса. После связывания ионов с насосом они погружаются внутрь его молекулы. Фосфорилирование насоса с помощью АТФ вызывает «закупорку» ионов натрия (Na3Ei-P). Переход к конформации Е2 открывает «карман» во внеклеточную среду. После связывания двух ионов калия процесс повторяется в обратной последовательности, только ионы калия выводятся из клетки.

| АТФаза |
| Рис. 2.4.Первично активный транспорт ионов кальция через мембрану. |
| Мембрана саркоплазматического ретикулума |
| Внешняя сторона АТФ — ► АДФ+Ф |
| + + + 2 Са++ Внутренняя сторона |

канала, поэтому во время транспорта ионов Na + и К + через мембрану клетки они окружены молекулой насоса и не могут взаимодействовать с другими ионами. За один цикл активности насоса три иона натрия выводятся из клетки, а внутрь транспортируются два иона калия.
Транспорт ионов Са 2+ через мембрану. В мембране саркоплазматического ретикулума всех типов мышечных клеток локализована Са 2+ — АТФаза (рис. 2.4).Функция этого насоса заключается в поддержании низкой концентрации ионов кальция в цитоплазме мышечных клеток за счет депонирования ионов в саркоплазматический ретикулум. Поддержание низкой концентрации ионов Са 2+ в саркоплазме является основным условием расслабления мышечных клеток (менее 10“ 7 моль/л, против 10“ 3 —10″ 2 моль/л в ретикулуме). Са ++ -АТФ-аза функционирует следующим образом. В присутствии АТФ после присоединения со стороны цитоплазмы иона кальция к кальций-связывающему участку Са 2+ -АТФаза фермент изменяет свою конформацию, и область связывания ионов Са 2+ оказывается внутри саркоплазматического ретикулума. При этом сродство ионов к Са 2+ -АТФазе уменьшается, и ионы Са 2+ высвобождаются во внутриретикулярное пространство. Под действием ионов Mg 2+ саркоплазматического ретикулума фермент Са 2+ -АТФаза дефосфорилируется и кальций-связывающий участок вновь оказывается снаружи мембраны. В такой последовательности повторяется цикл работы кальциевого насоса. В саркоплазме мышечных клеток ионы кальция принимают участие в механизме мышечного сокращения либо выполняют функцию вторичного посредника (описаны ниже).
Читайте также: Кенгуру из ткани своими руками
Транспорт протонов через мембрану. Протонная помпа (Н + -АТФаза) транспортирует протоны через внутренние мембраны митохондрий. Протонная помпа в митохондриях увеличивает градиент электрохимического потенциала ионов водорода на мембране до порогового или критического уровня, который необходим для синтеза АТФ. Протонная помпа увеличивает электрическую и концентрационную составляющую электрохимического потенциала на мембране митохондрий.
2.1. 1.5. Вторично-активный транспорт
В возбудимых клетках имеет место активный транспорт веществ через мембрану клеток против их концентрационного градиента, если эквивалентное количество энергии высвобождается в результате движения другого вещества по его концентрационному градиенту. Этот вид трансмембранного транспорта называется вторично активным. Вторично активный транспорт называется также сопряженным транспортом, поскольку более одного вещества транспортируется через мембрану клетки с помощью интегрального белка-переносчика. При этом виде транспорта молекулы веществ конкурируют между собой за белок-переносчик. Например, если ве-
щество А равномерно распределено по обе стороны мембраны, а вещество Б создает более высокую концентрацию по одну стороны мембраны, то конкуренция вещества Б за переносчик по сравнению с веществом А будет тем больше, чем выше будет градиент концентрации вещества Б. При этом, если белок-переносчик переносит две субстанции, то это называется котранспортом. Если два вещества переносятся через мембрану одновременно в одном направлении, то это называется симпортом. Если вещества переносятся через мембрану одновременно, но в противоположных направлениях, это называется антипортом. Как правило, натрий-калиевый насос создает градиент концентрации ионов натрия и калия по обе стороны от мембраны клеток, который обеспечивает сопряженный или вторично активный транспорт.
Этот вид активного транспорта широко представлен в физиологических процессах различных органов. В скелетных мышцах, кардиомиоцитах и гладкомышечных клетках с функцией Nа + /К + -АТФазы связан транспорт ионов Са 2+ через плазматическую мембрану при участии Na*/Са 2+ -обменника. Этот белок транспортирует ионы Са 2+ через мембрану клеток в обмен на ионы натрия и за счет энергии На + /К + -АТФазы. Кроме того, в мембранах клеток имеется белок, обменивающий внеклеточные ионы натрия на внутриклеточные протоны, — N а*/Н + -обменник, который поддерживает постоянство pH внутриклеточной среды. Скорость обмена ионов при участии Na + /Ca 2+ — и Ма + /Н + -обменников пропорциональна электрохимическому градиенту ионов Na + по обе стороны мембраны. В невозбудимых клетках, например, слизистой оболочки тонкой кишки глюкоза транспортируется в эпителиоциты лишь в том случае, если ионы Na + одновременно с глюкозой связывается с белком-переносчиком.
2.1.1.6. Эндоцитоз и экзоцитоз
Молекулы некоторых веществ, например химических передатчиков возбуждения в нервной системе (медиаторов) или гормонов, не способны проходит через мембрану клеток. Поэтому их движение через клеточную мембрану осуществляется путем эндоцитоза и экзоцитоза. При эндоцитозе субстанции первоначально адсорбируются специальными рецепторами на внешней стороне мембраны. С помощью мембранного белка клатрина, расположенного на внутренней поверхности мембраны клетки, в мембране формируется углубление, а затем образуется везикула и везикулярный комплекс «рецептор—клатрин—белок» оказывается внутри клетки. Комплекс утрачивает рецептор, белок клатрин вновь возвращается на внутреннюю поверхность клеточной мембраны, а поступивший внутрь белок метаболизируется в клетке.
Путем экзоцитоза из клетки выводятся крупные белковые молекулы, которые инкапсулируются мембранами эндоплазматического ретикулума в форме везикул. Подобные везикулы, а для многих клеток они представляют собой секреторные гранулы, транспортируются вдоль внутриклеточной микротрубулярной системы к мембране клеток, сливаются с клеточной мембраной в строго определенных местах мембраны, содержащей специальный белок синапсин. Процесс слияния секреторных гранул активируется ионами Са ++ . Мембрана секреторной везикулы соединяется с мембраной клетки, и секретируемые субстанции оказываются во внеклеточной среде.
2.7.7.7. Внутриклеточный транспорт молекул

| Аксон Сома нейрона ◄———— |
| Рис. 2.5. Внутриклеточное движение органелл с помощью белков, связанных с микротрубочками: динеин перемещает органеллы нейронов в направлении их сомы, а кинезин — в направлении нервной терминали. |
различные субстанции также транспортируются с помощью специальных белков. Например, в нервных клетках имеет место аксонный транспорт везикул как в направлении нервных окончаний (антероградно), так и в направлении тела нейронов (ретроградно). Внутриклеточные транспортные процессы осуществляются вдоль цитоплазматических
микротрубочек при участии
специфических белков кинезина и динеина (рис. 2.5).При этом кинезин транспортирует везикулы по микротрубочкам аксона в направлении терминалей нейронов, а динеин — в направлении их сомы.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом
- Правообладателям
- Политика конфиденциальности
Мастерица © 2023
Информация, опубликованная на сайте, носит исключительно ознакомительный характер