
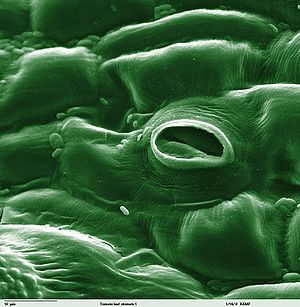
У́стьице (лат. stoma , от греч. στόμα — «рот, уста») — в ботанике это по́ра, находящаяся на нижнем или верхнем слое эпидермиса листа растения, через которую происходит испарение воды и газообмен с окружающей средой.
История исследований
Хотя учёные давно знали об испарении воды поверхностью листа, первым, кто наблюдал устьица, был итальянский натуралист Марчелло Мальпиги, который это открытие опубликовал в 1675 году в своей работе Anatome plantarum. Однако он не понял их настоящую функцию. В то же время его современник Неемия Грю развил гипотезу об участии устьиц в вентиляции внутренней среды растения и сравнил их с трахеями насекомых. Прогресс в изучении наступил в XIX веке, и тогда же, в 1827 году, швейцарским ботаником Декандолем было впервые использовано слово „stoma“. Изучением устьиц в то время занимались Гуго фон Моль, который открыл основной принцип открывания устьиц и Симон Швенденер, классифицировавший устьица по типу их конструкции.
Некоторые аспекты функционирования устьиц продолжают интенсивно изучаться и в настоящее время; материалом в основном служат Коммелина обыкновенная ( Commelina communis ), Боб садовый ( Vicia faba ), Кукуруза сахарная ( Zea mays ). [1]
Строение
Размеры устьица (длина) колеблются в пределах 0,01–0,06 мм (крупнее бывают устьица полиплоидных растений и у листьев, растущих в тени. Самые крупные устьица были обнаружены у вымершего растения Zosterophyllum, 0,12 mm (120 µm) [1] Пора состоит из пары специализированных клеток, называемых замыкающими (cellulae claudentes), которые регулируют степень открытости поры, между ними располагается устьичная щель (porus stomatalis). Стенки замыкающих клеток утолщены неравномерно: направленные к щели (брюшные) толще стенок, направленных от щели (спинных). Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Когда воды мало, замыкающие клетки плотно прилегают друг к другу и устьичная щель закрыта. Когда воды в замыкающих клетках много, то она давит на стенки и более тонкие стенки растягиваются сильнее, а более толстые втягиваются внутрь, между замыкающими клетками появляется щель [2] . Под щелью расположена подустьичная (воздушная) полость, окружённая клетками мякоти листа, через которую непосредственно и происходит газообмен. Воздух, содержащий диоксид углерода (углекислый газ) и кислород, проникает внутрь ткани листа через эти поры, и далее используется в процессе фотосинтеза и дыхании. Избыточный кислород, произведённый в процессе фотосинтеза внутренними клетками листа, выходит обратно в окружающую среду через эти же поры. Также, в процессе испарения через поры выделяются пары воды. Клетки эпидермиса, примыкающие к замыкающим, получили название сопровождающих (побочных, соседних, околоустьичных). Они участвуют в движении замыкающих клеток. Замыкающие и сопровождающие клетки образуют устьичный комплекс (устьичный аппарат). Наличие или отсутствие устьиц (видимые части устьиц называют устьичными линиями) часто используют при классификации растений.
Типы устьиц
Число сопровождающих клеток и их расположение относительно устьичной щели позволяют выделить ряд типов устьиц:
- аномоцитный — сопровождающие клетки не отличаются от остальных клеток эпидермиса, тип весьма обычен для всех групп высших растений, за исключением хвойных;
- диацитный — характеризуется только двумя сопровождающими клетками, общая стенка которых находится под прямым углом к замыкающим клеткам;
- парацитный — сопровождающие клетки располагаются параллельно замыкающим и устьичной щели;
- анизоцитный — замыкающие клетки окружены тремя сопровождающими, одна из которых заметно крупнее или мельче остальных, такой тип обнаружен только у цветковых растений;
- тетрацитный — четыре сопровождающие клетки, характерен для однодольных;
- энциклоцитный — сопровождающие клетки образуют узкое колесо вокруг замыкающих клеток;
- актиноцитный — несколько сопровождающих клеток, радиально расходящихся от замыкающих клеток;
- перицитный — замыкающие клетки окружены одной побочной сопровождающей клеткой, устьице не соединено с сопровождающей клеткой антиклинальной клеточной стенкой;
- десмоцитный — замыкающие клетки окружены одной сопровождающей клеткой, устьице соединено с ней антиклинальной клеточной стенкой;
- полоцитный — замыкающие клетки окружены одной сопровождающей не полностью: к одному из устьичных полюсов примыкает одна или две эпидермальные клетки; устьице прикреплено к дистальной стороне единственной сопровождающей клетки, имеющей U-образную или подковообразную форму;
- стефаноцитный — устьице, окружённое четырьмя или более (обычно пять-семь) слабодифференцированными сопровождающими клетками, образующими более или менее отчётливую розетку;
- латероцитный — такой тип устьичного аппарата рассматривается большинством ботаников как простая модификация аномоцитного типа.
У двудольных распространённым является парацитный тип устьиц. Замыкающие клетки почковидной (бобовидной) формы — такими они видны с поверхности листа — несут хлоропласты, тонкие неутолщённые участки оболочки образуют выступы (носики) закрывающие устьичную щель.
Наружные стенки замыкающих клеток обычно имеют выросты, что хорошо видно на поперечном разрезе устьица. Пространство, ограниченное этими выростами, называют передним двориком. Нередко аналогичные выросты наблюдаются и у внутренних оболочек замыкающих клеток. Они образуют задний дворик, или внутренний, соединённый с крупным межклетником — подустьичной полостью.
У однодольных парацитное строение устьиц отмечено у злаковых. Замыкающие клетки имеют гантелевидную форму — сужены в средней части и расширены на обоих концах, при этом стенки расширенных участков очень тонкие, а в средней части замыкающих клеток сильно утолщены. Хлоропласты располагаются в пузыревидных окончаниях клеток.
Движение замыкающих клеток
Механизм движения замыкающих клеток весьма сложен и неодинаков у разных видов. У большинства растений при неодинаковом водоснабжении в ночные часы, а иногда и днём тургор в замыкающих клетках понижается, и устьичная щель замыкается, снижая тем самым уровень транспирации. С повышением тургора устьица открываются. Считают, что главная роль в изменении тургора принадлежит ионам калия. Существенное значение в регуляции тургора имеет присутствие в замыкающих клетках хлоропластов. Первичный крахмал хлоропластов, превращаясь в сахар, повышает концентрацию клеточного сока. Это способствует притоку воды из соседних клеток и повышению тургорного давления в замыкающих клетках.
Расположение устьиц
Двудольные растения, как правило, в нижней части листа имеют больше устьиц, чем в верхней. Это объясняется тем, что верхняя часть горизонтально-расположенного листа, как правило, лучше освещена, и меньшее количество устьиц в ней препятствует избыточному испарению воды. Листья с устьицами, расположенными на нижней стороне, называются гипостоматическими.
Читайте также: Ткань с углеродной нитью
У однодольных растений наличие устьиц в верхней и нижней части листа различно. Очень часто листья однодольных растений расположены вертикально, и в этом случае количество устьиц на обоих частях листа может быть одинаково. Такие листья называются амфистоматическими.
У плавающих листьев на нижней части листа устьица отсутствуют, так как они могут впитывать воду через кутикулу. Листья с устьицами, расположенными на верхней стороне, называются эпистоматическими. У подводных листьев устьица отсутствуют совсем.
Устьица хвойных растений обычно спрятаны глубоко под эндодермой, что позволяет сильно снизить расход воды зимой на испарение, а летом — во время засухи.
У мхов (исключение антоцеротовые)настоящие устьица отсутствуют.
Устьица также различаются по уровню расположения относительно поверхности эпидермиса. Некоторые из них расположены вровень с другими эпидермальными клетками, другие подняты выше или погружены ниже поверхности. У однодольных, листья которых растут преимущественно в длину, устьица образуют правильные параллельные ряды, тогда как у двудольных они располагаются беспорядочно.
Углекислый газ
Так как углекислый газ является одним из ключевых реагентов в процессе фотосинтеза, у большинства растений устьица в дневное время открыты. Проблема состоит в том, что при входе воздух смешивается с парами воды, испаряющимися из листа, и поэтому растение не может получить углекислый газ, одновременно не потеряв некоторое количество воды. У многих растений существует защита от испарения воды в виде закупоривающих устьица восковых отложений.
Пара замыкающих клеток с устьичной щелью называют устьице ткань кожица
Кожица, или эпидермис. Кожица , или эпидермис , представляет собой первичную ткань листьев и стеблей. Кожица состоит из плотно сомкнутых клеток, имеющих в плане у большинства растений более или менее извилистые очертания (рис. 86). На поперечных разрезах органа клетки кожицы обычно имеют четырехугольные или пятиугольные очертания. Иногда наружная и внутренняя стенки слегка выпуклы. Клетки кожицы органов, вытянутых в длину (листовых черешков, листовых пластинок линейного и ланцетного очертания, стеблей), обычно сильно удлинены в направлении, параллельном продольной оси органа. У некоторых растений, например у многих злаков, кожица состоит из клеток нескольких типов (рис. 87).
Оболочка эпидермальных клеток утолщается обычно неравномерно: в каждой клетке наиболее толста наружная стенка, боковые стенки несколько тоньше, внутренняя — сравнительно тонка. Встречаются исключения из этого правила. Боковые и внутренние стенки клеток обычно имеют поры (рис. 87), наружная же стенка бывает снабжена ими в весьма редких случаях. Оболочка клеток состоит в основном из целлюлозы. У многих растений (особенно у злаков, осоковых, хвощей) наружная стенка кожицы пропитана кремнеземом; у некоторых растений в наружной стенке, реже в боковых (у драцен) и даже во внутренних откладываются мелкие кристаллы или зернышки щавелевокислого кальция.
Поверхность кожицы покрыта пленкой — кутикулой (надкожицей), состоящей из кутина. Кутикула очень тонка или вовсе отсутствует на поверхности конусов нарастания. Часто кутикула имеет гладкую поверхность, в иных случаях образует выступы в виде бугорков, перекладин, прямых или волнистых ребер и т. д. Эти выступы состоят нацело из

Рис. 86. Участки кожицы листа лютика ползучего (Ranunculus repens):
1 — с верхней стороны листа; 2 — с нижней его стороны.
кутикулы или же являются выступами целлюлозной стенки, покрытыми кутикулой.
На поперечных срезах кожицы иногда видно, что кутикула пронизывает и часть толщи стенок. Кутикула сильно преломляет свет и поэтому хорошо видна на срезах. Изолированная кутикула обычно представляет бесцветную бесструктурную сплошную пленку. У некоторых растений (например, у золотого дерева Aucuba) кутикула состоит из пластинок, каждая из которых принадлежит одной эпидермальной клетке; над пограничными участками клеток кожицы кутикулы нет или она очень тонка.
Мощность кутикулы зависит от вида и возраста растения и условий местообитания (рис.88); на поверхности стеблей с многолетней кожицей (у некоторых видов калины, клена) кутикула разрывается и подновляется изнутри путем образования новых ее наслоений. Кутикула листьев некоторых осоковых подвергается окремнению.
У многих растений во внешних стенках клеток кожицы, иногда в боковых, еще реже во внутренних, образуются кутикулярные слои — прослойки в целлюлозной клеточной оболочке, содержащие кутин (рис. 76). Кутикула и кутикулярные слои нередко пропитаны воском; при подогревании препаратов в воде под покровным стеклом он выплавляется в виде капель. У некоторых растений воск покрывает кутикулу в виде мелких зерен, расположенных равномерным слоем (на листьях и стеблях многих злаков, лилейных, например тюльпанов), тонких палочек, часто изогнутых и на конце закрученных (на стеблях некоторых злаков, сахарного тростника, рис. 89), сплошных корочек, тонких — 1μ толщиной (на листьях туй, молодил).
Восковой налет, как и кутикула, снижает транспирацию

Рис. 87. Часть кожицы листа пшеницы, в плане:
1 — замыкающие клетки устьиц; 2 — побочные клетки; 3 — клетки эпидермиса с многочисленными порами; в полости клеток — постенная протоплазма и ядро, в некоторых из клеток не зарисованные.

Рис. 88. Часть поперечного разреза через листообразный бугорок кактуса Ariocarpus retusus; чрезвычайно толстая кутикула пронизана фигурными ходами над устьицами. Под кожицей — зеленая паренхима.
органов. Воск делает поверхность органов несмачиваемой: с них быстро стекает вода, чем предупреждается капиллярное закупоривание водой устьиц и заселение поверхности растений мелкими эпифитами.

Рис. 89. Часть поперечного разреза через узел стебля сахарного тростника:
к — кожица: в — восковые палочки.
Поэтому понятно, почему стебли и листья многих подводных растений лишены кутикулы.
Клетки кожицы содержат тонкий постенный слой протоплазмы с ядром и пластидами и крупную центральную вакуолю. Обычно из пластид здесь имеются лейкопласты.
У многих растений в вакуолях клеток кожицы — нередко только на нижней стороне листа — содержится антоциан.
Читайте также: Ткань растягивается больше по направлению чего
В вакуолях эпидермальных клеток нередко встречаются дубильные вещества, соли щавелевой кислоты в виде растворов (у щавелей, кислиц) или кристаллов; реже здесь имеются алкалоиды, ядовитые глюкозиды, гипс (у каперсов — в виде кристаллов), кальциевые соли яблочной и винно-каменной кислот (у винограда).
Хлоропласты и хромопласты в эпидермисе цветковых растений встречаются очень редко, только у некоторых теневых и водных растений 1 .
Устьица. Для осуществления газообмена между внутренними тканями растений и внешней средой и для транспирации (испарения воды) в кожице имеются устьица.
Устьице состоит из двух специализированных замыкающих клеток и щелевидного отверстия между ними — устьичной щели . При рассматривании кусочка кожицы, положенного плашмя на предметное стекло, устьице обычно видно как пара бобовидных или полулунных клеток, обращенных друг к другу вогнутыми сторонами (рис. 90-92). Оболочка каждой замыкающей клетки на внутренней стороне сильно и неравномерно утолщена, на противоположной, наружной, стороне — тонка.
На поперечном разрезе, проведенном через середину устьичной щели перпендикулярно к ней (рис. 91), видны выросты стенок замыкающих клеток устьица, называемые отрогами или клювиками.
Щель устьица имеет сложное очертание: промежуток между наружными отрогами ведет в преддверие, или передний дворик устьица . Дальше идет наиболее узкая часть устьичной щели — центральная щель устьица , находящаяся между тонкими участками внутренних стенок. Центральная щель расширяется дальше в задний дворик . Отверстие между отрогами нижних стенок замыкающих клеток

Рис. 90. Кожица листа гиацинта (Hyacinthus orientalis) с устьицами, в плане:
з — замыкающие клетки, о — щель устьица.

Рис. 91. Устьице с окружающими клетками эпидермиса (листья тимьяна Thymus):
1 — в плане; 2 — в разрезе; щ — центральная щель устьица; а — тонкая стенка замыкающей клетки устьица, обращенная к соседним клеткам кожицы; в замыкающих клетках видно ядро (я) с ядрышком и хлорофилловые зерна (x); к — кутикула; м — межклетная воздухоносная полость («дыхательная полость»).

Рис. 92. Устьичный аппарат листа свеклы (Beta vulgaris):
1, 3 — устьице в плане; 2, 4 — в разрезе; 1, 2 — устьице закрыто; 3, 4 — устьице открыто.
сообщает задний дворик с дыхательной полостью (рис. 91, 2; 93).
Возвращаясь к устьицу в плане (рис. 91, 1), отметим, что щель устьица в оптических сечениях, параллельных поверхности эпидермиса, имеет форму узкой двояковыпуклой линзы; в плане обычно видно очертание щели на уровне наружного отверстия и на уровне середины центральной щели. Наружные стенки, а у некоторых растений отчасти также и внутренние покрыты кутикулой; оторочки двориков иногда состоят сплошь из кутикулы. Функционирующие замыкающие клетки живые; они содержат постенный слой протоплазмы с ядром и с хлорофилловыми зернами.
Обычно устьица распределяются на поверхности листа или стебля поодиночке и более или менее равномерно (рис. 90). У некоторых растений (например, у злаков) они располагаются продольными рядами, а у других (у олеандра, селезеночника) — группами.
Образование замыкающих клеток устьиц происходит при неравномерном делении меристематических клеток эпидермиса, в результате чего возникают инициальные клетки устьиц. В простейших случаях инициальная клетка становится производящей клеткой устьица. Производящая клетка образует путем деления пару замыкающих клеток устьица; расщепление перегородки между ними ведет к образованию устьичной щели. У некоторых растений инициальная клетка образует путем деления целые группы клеток — пару замыкающих и две или несколько побочных клеток

Рис. 93. Часть поперечного разреза листа Aloe disticha с устьицем, погруженным в ткань листа:
э — кожица с толстыми наружными стенками, покрытыми мощной кутикулой; у — полость над устьицем; з — замыкающие клетки с кутикулярными выступами (обозначенными черным цветом); м — воздухоносный межклетник («дыхательная полость»); х — клетки хлорофиллоносной паренхимы.
устьица; последние отличаются от прочих клеток кожицы меньшей величиной и своеобразными очертаниями (рис. 94).
Устьичный аппарат регулирует газообмен растения. Замыкающие клетки могут смещаться, изменять объем и форму, в результате чего меняется и очертание устьичной щели: она может быть более или менее широко открыта или же вовсе закрыта (рис. 92). При открывании и закрывании устьиц происходят изменения живого содержимого замыкающих клеток. При открытой щели устьица протоплазма в них более вязка, чем при закрытой. Ядро изменяет форму: у кормовых бобов при длительно открытом устьице оно становится веретеновидньш, при закрытом — округло-лопастным, у георгина — наоборот (рис. 95).
Открывание и закрывание устьиц обусловливается тургорными явлениями. Изменения тургора замыкающих клеток могут быть непосредственным следствием подвядания листа или повышения в нем содержания воды.
При повышении тургора замыкающих клеток объем их клеточной полости увеличивается, в результате чего стенки отдаляются одна от другой и щель открывается. При понижении тургора замыкающих клеток происходят изменения обратного рода: стенки сближаются до соприкосновения и щель устьица замыкается.
Изменения тургора в замыкающих клетках регулируются превращением в них крахмала хлоропластов в сахар, и обратно — сахара в крахмал: при осахаривании крахмала в замыкающих клетках повышается концентрация клеточного сока и увеличивается их сосущая сила. В результате замыкающие клетки насасывают воду (частично за счет соседних клеток), что вызывает изменение их объема и раскрывание устьичной щели (рис. 96). Превращение сахара замыкающих клеток в крахмал влечет аналогичным образом закрывание щели устьица.
Косвенное участие в закрывании и открывании устьиц могут принимать не только клетки, соседние с замыкающими, но и более дальние побочные клетки устьиц и рядовые клетки кожицы. При продолжительном дожде все клетки кожицы насыщаются водой, тургор их сильно повышается и замыкающие клетки устьиц сдавливаются; в результате устьичные щели пассивно замыкаются. Через некоторое время после прекращения дождя нормальная работа устьиц восстанавливается.
Тургорная теория действия устьиц, выдвинутая Молем в 1856 г., подверглась дополнениям со стороны ряда ученых, указывающих на возможное значение изменений проницаемости протоплазмы замыкающих клеток.
Читайте также: Структурные элементы входящие в состав тканей
В устройстве устьиц встречается большое разнообразие. Форма щели может быть значительно упрощена или усложнена по сравнению с вышеописанной, типичной. Наиболее упрощена она у растений с листьями, плавающими на воде (у белой кувшинки): щель имеет вид воронки, широким концом соединяющейся с дыхательной полостью, а узким переходящей в наружное отверстие; степень раскрытия щели устьица зависит от состояния этого отверстия.
У растений с листьями, расположенными более или менее горизонтально, особенно же у деревьев и у трав тенистых местообитаний, устьица расположены преимущественно или исключительно на их нижней стороне.
У многих растений, преимущественно у приуроченных к сухим, солнечным и ветреным местообитаниям, устьица погружены в глубь органа за счет сильно кутинизированных выступов соседних клеток (рис. 93). Иногда устьица располагаются целыми группами в глубине ямочек (как у олеандра). У некоторых видов австралийских бэнксий (Banksia) на дне и вокруг ямок с устьицами имеются длинные белые волоски; образуются, таким образом, убежища с затишьем от ветра и затенением от солнца.
Устьица имеют представители всех классов высших растений 2 . У незеленых паразитов и сапрофитов устьица малочисленны либо редуцированы или же вовсе не образуются. Подземные листовые и стеблевые органы весьма бедны устьицами. У водных растений устьиц сравнительно мало; у многих из них устьица редуцированы. Устьица можно отнести к системе тканей проветривания, но обычно их изучают вместе с кожицей.

Рис. 94. Развитие устьичного аппарата в листе кукурузы, в плане:
А — три производящие клетки устьиц (1, 1, 1) уже отделились перегородками от инициальных клеток; в соседних клетках происходит деление ядра (2, 2), предшествующее образованию побочной клетки; Б — при каждой из производящих клеток трех устьиц образовались две побочные клетки (3); В — в трех производящих клетках устьиц происходит деление ядра (4, 4), предшествующее образованию замыкающих клеток; Г — производящая клетка поделилась на две клетки, дифференцирующиеся в пару замыкающих клеток устьица (5, 5), с устьичной щелью между ними; 6, 6 — побочные клетки; Д — сформированный устьичный аппарат, состоящий из двух замыкающих клеток (5, 5) и двух побочных клеток (6, 6).
Волоски (трихомы). У большинства высших растений некоторые или многие из клеток кожицы образуют выросты, так называемые волоски , имеющие различную форму, сосочков, бугорков, щетинок, кустиков, звездчатых чешуек и т. д. В простейшем случае волосок представляет сосочек — короткий округло-конический вырост клетки кожицы. Сосочками же

Рис. 95. Изменение ядра и крахмальных зерен в замыкающих клетках устьица листа георгина:
а — устьице открыто, в замыкающих клетках очень мелкие крахмальные зерна, ядро округло-амебоидного очертания, с вакуолей в центре; б — устьице закрыто, в замыкающих клетках — сравнительно крупные зерна крахмала, клеточное ядро веретенообразно, с крупными ядрышками в центре.
являются волоски почти всех типов в ранней стадии их образования. Встречаются волоски, не отделяющиеся перегородкой от произведшей их клетки кожицы, как например щетинки многих бурачниковых. В большинстве случаев волосок отделяется перегородкой от произведшей его эпидермальной клетки — «материнской клетки волоска». Отчленяющийся волосок редко остается одноклеточным; чаще он становится многоклеточным. Многоклеточные волоски очень разнообразны. Среди них различают 1) линейные, состоящие из клеток, расположенных в ряд (листья картофеля); 2) кустисто-ветвистые (коровяки, рис. 97, 6); 3) чешуйчатые и звездчато-чешуйчатые (Elaeagnus, рис. 97, 7, 8) и 4) массивные, представляющие как бы пучок спаявшихся нитевидных волосков (волоски многих губоцветных). Содержимое волосков некоторых категорий во взрослой стадии разрушается, полости их обычно заполняются воздухом. Такие волоски, имеющие обычно белый, либо сероватый, или желтоватый цвет, можно считать одним из приспособлений, умеряющих действие на лист или стебель горячих лучей солнца и ослабляющих иссушающее действие ветров.
Некоторые волоски могут служить растению защитой от поедания животными; так, тесно расположенные жесткие, острые щетинки и войлочный покров предохраняют листья и стебли от объедания слизняками и отчасти от поедания травоядными млекопитающими.
Жгучие волоски . Жгучий волосок крапивы — крупная живая клетка с пузыревидным основанием, сидящим в чашевидной многоклеточной «подставке». Волосок представляет в миниатюре как бы полую иглу шприца и заканчивается косо поставленной головкой. Так как оболочка волоска, богатая известью и кремнеземом, очень ломка и к тому же в месте перехода «иглы» в «головку» весьма тонка, то при малейшем прикосновении животного или человека к головке последняя отламывается, острый край «иглы» вонзается в кожу, и клеточный сок волоска, содержащий жгучие вещества, впрыскивается в нее (рис. 97, 9-12).
Прикосновение руки к листьям некоторых крапив тропической флоры (например, Urtica urentissima Индонезии) вызывает болезненное или даже опасное для жизни воспаление.
Некоторые категории волосков относятся не к покровной, а к иным тканям (см. ниже).
К покровным тканям относятся еще перидерма и корка (описание их см. на стр. 173-175).

Рис. 96. Схематизированный оптический разрез устьица зимовника (Heiieborus sp.) в закрытом и открытом состоянии.
Наружные очертания и клеточная полость замыкающих клеток показаны при закрытой щели устьица и при открытой щели. Клеточная полость обеих замыкающих клеток при закрытом устьице заштрихована.

Рис. 97. Различные формы волосков:
1, 2 — подмаренник (Сalium); 3 — хмель (Humulus); 4 — наперстянка (Digitalis); 5 — камнеломка (Saxifraga); 6 — коровяк (Verbascum); 7, 8 — лох (Elaeagnus); 9-12 — жгучий волосок крапивы (Urtica); а — обламывающаяся верхушка волоска.
1 Наличие хлоропластов характерно для эпидермиса споровых растений. Сравнительно крупные и ярко окрашенные хлорофилловые зерна содержат клетки кожицы листьев папоротников.
2 Однако одна из групп печеночников — маршанциевые — лишена настоящих устьиц: слоевище их имеет «дыхательные отверстия», аналогичные по функции устьицам, но совершенно отличные по устройству (см. в разделе «Систематика растений»).
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом