
Электропроводность в неживой природе. Электропроводность живых тканей. Диэлектрические свойства живых тканей. Физические поля биологических объектов
Страницы работы

ПАССИВНЫЕ ЭЛЕКТРИЧЕСКИЕ СВОЙСТВА ЖИВЫХ ТКАНЕЙ.
Основными пассивными электрическими характеристиками вещества являются: диэлектрическая проницаемость, удельная электрическая проводимость. Эти величины и связанные с ними электроемкость, импеданс существенно зависят от состояния организма и активно используются в диагностике
1) ЭЛЕКТРОПРОВОДНОСТЬ В НЕЖИВОЙ ПРИРОДЕ
Пассивные электрические свойства, веществ в неживой природе хорошо изучены. Переходя к изучению свойств живых тканей (они значительно сложнее) мы будем опираться на их неживые модели.
Электропроводность — это величина, обратная электрическому сопротивлению G=1/R
Она измеряется в сименсах: 1 См = 1 Ом -1
Удельная электрическая проводимость
Для проводников Λ > 10 6 См м -1
Для диэлектриков Λ -8 См м -1 .
Закон Ома можно переписать через плотность тока, удельную электрическую проводимость и напряженность электрического поля
Рассмотрим электропроводность в неживой природе. В зависимости от вида и природы носителей проводимость бывает электронной, ионной и дырочной. Электронной проводимостью обладают металлы. Жидкие электролиты обладают ионной проводимостью. Дырочная проводимость образуется при разрыве валентной связи, приводящей к появлению вакантного места для электрона. Полупроводники обладают электронной и дырочной проводимостью.
Электрическая проводимость прямо и косвенно зависит от систем атомов и молекул. Она обусловлена положением электронов относительно ядра, их спинами и магнитными моментами. В металлах некоторые электроны утрачивают связь со своими атомами и могут свободно передвигаться. В растворах основными носителями зарядов являются ионы. Прохождение тока через растворы вызывает перемещение вещества в нем, и. следовательно, изменение его химических свойств. Этим растворы отличаются от металлических проводников.
Современные представления об электрических свойствах живых тканей основаны на фактах о молекулярной организации мембран и, в значительно меньшей степени, на сведениях о квантовомеханических свойствах физиологически активных молекул.
Живые ткани являются композиционными средами, т.к. одни структурные элементы являются проводниками, а другие диэлектриками. Кроме того, некоторые макромолекулы обнаруживают полупроводниковые свойства (А.Сцент-Дъердьи).
Найдены элементы и даже молекулы, играющие роль доноров и акцепторов, + крайне малые значения энергии связи электронов в биологических молекулах.
Электропроводность живых тканей определяется в основном злектрическими свойствами лимфы, крови и межклеточной жидкости. Для них Λ ≈ 1 См-м -1 . Минимальная Λ у костной ткани, порядка 10 -7 См-м -1 . Однако, Λ целых органов на 4 — 6 порядков ниже L жидкостей, находящихся в них. Причина в том, что электролиты занимают малые объемы. Так, в клетках электролиты заключены в мельчайшие отсеки — «компартменты», образованные биомембранами, составляющими более 50% массы клетки. Мембраны являются диэлектриками.
Для диагностики важно отметить, что наблюдается различие в проводимости опухолей и нормальной ткани, а так же зависимость проводимости от насыщенности тканей кислородом.
1). При измерении пассивных электрических характеристик живых тканей через них необходимо пропускать электрический ток. что приводит к раздражению тканей и изменению их свойств. Результаты измерений зависят от силы тока, его типа, плотности тока на электродах.
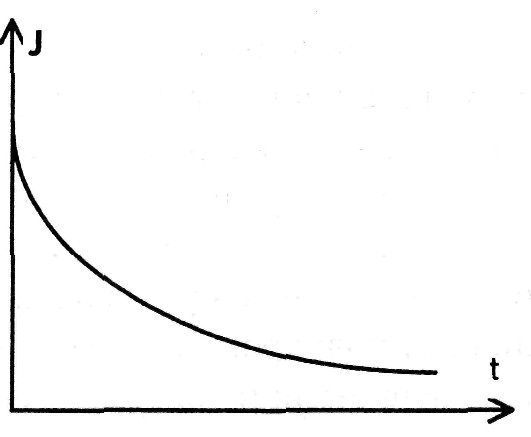
При прохождении постоянного тока через ткани его первоначальная сила не остается постоянной, а падает с течением времени до определенного уровня (рис.).

Это означает, что происходит уменьшение проводимости живой ткани. Процесс установления равновесия (релаксационный процесс) имеет сложную природу, можно предложить лишь некоторые его модели:
Поляризация — это процесс перемещения связанных зарядов под действием электрического поля и образование вследствие этого электродвижущей силы, которая направлена противоположно внешнему полю. Ее называют ЭДС поляризации. Для живой ткани закон Ома можно записать в виде:
где U — приложенное напряжение, I — сила тока, R – активное сопротивление ткани, εп — ЭДС поляризации, зависящая от U и времени.
б) процесс заряда тканевых емкостей.
2). Измерения проводимости в БАТ (метод Фолля). Оставляя в стороне достоверность этой диагностики, рассмотрим возможные причины неоднозначности подобных измерений:
а) Исследования проводятся по точкам, расположенным в направлении «меридианов», которые были интуитивно установлены в восточной медицине. Недавно было обнаружено, что в этих направлениях между клетками тканей имеется повышенное число щелевых контактов, проницаемых для ионов, чем и объясняется большая проводимость.
б) При измерениях используют маленькие по площади электроды, и от величины силы, с которой электрод прижимается к коже, существенно зависят получаемые результаты.
в) Ток от внешнего источника так же изменяет состояние БАТ.
г) проводимость меридианов зависит не только от патологий соответствующих органов, но и от индивидуальных биоритмов человека.
3) ДИЭЛЕКТРИЧЕСКИЕ СВОЙСТВА ЖИВЫХ ТКАНЕЙ.
Читайте также: Как по английски будет торговля тканями
Диэлектрические свойства биологических тканей обусловлены явлениями поляризации и компартментной структурой.
Диэлектрическая проницаемость где Ei — напряженность внутреннего поля.
Электрическое поле внутри тканей меньше внешнего так как поляризованные и ориентированные определенным образом атомы, молекулы и компартменты создают электрическое поле Еп направленное против внешнего поля (рис.).
Рассмотрим виды поляризации характерные дляживой ткани.
1) Электронная поляризация — смещение электронов на своих орбитах относительно положительно заряженных ядер в атомах и ионах. В результате такого смещения атом или ион превращаются в индуцированный диполь. Время релаксации (установления или исчезновения) электронной поляризации равняется 10 -16 —10 -14 с

2) Дипольная (ориентационная) поляризация— ориентация полярных молекул под действием внешнего электрического поля. Дипольная поляризация преобладает в таких веществах как спирт и вода. Молекулы белков и других высокомолекулярных соединений вследствие диссоциации моногенных групп, а также вследствие адсорбции ионов обладают значительными дипольными моментами. Время релаксации дипольной поляризации совпадает со временем поворота молекул и зависит от вязкости среды, температуры и радиуса молекул, поэтому оно изменяется в широких пределах от 10 -13 до 10 -7 с
3) Макроструктурная поляризация — перемещение свободных ионов и электронов в пределах компартментов. В результате этого компартменты приобретают дипольный момент и ведут себя как гигантские поляризованные молекулы. Время релаксации макроструктурной поляризации лежит в пределах от 10 -8 до 10 -3 с. Макроструктурная поляризация происходят во всем объеме клеток и играет основную роль в живых тканях, определяя их высокую диэлектрическую проницаемость в постоянном электрическом поле (ε≈ 10 4 ).
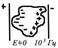
При включении электрического поля первыми возникают и исчезают при выключении те виды поляризации, которые имеют меньшее время релаксация. В переменном электрическом поле возникает зависимость диэлектрической проницаемости живых тканей от частоты поля — дисперсия (рис.3).
Для компартментов, полярных молекул (их дипольных моментов) существует максимальная частота внешнего ЭМП, обратная времени релаксации, которую они способны воспроизвести своими «поворотами» (изменением поляризации) в нем для компенсации внешнего поля. Высокая диэлектрическая проницаемость живых тканей на низких частотах обусловлена компартментами. При частотах внешнего поля до 10 3 Гц переориентация дипольных моментов компартментов отстает от изменения поля, и диэлектрическая проницаемость тканей падает, это область α-дисперсии (см. рис.3). На более высоких частотах (до 10 8 Гц) внешнее поле успевают частично компенсировать полярные макромолекулы (β-дисперсия), на сверхвысоких частотах (10 11 Гц ) — молекулы воды (γ- дисперсия).
В живых тканях под воздействием ЭМП возникают токи проводимости, обусловленные смещением (колебаниями) ионов и токи смещения, связанные с переориентацией дипольных моментов. Можно считать что в цепи переменного тока через конденсатор течет именно ток смещения, т.к. перемещения зарядов через конденсатор не происходит.
Активные и пассивные электрические свойства органов и тканей
Активные электрические свойства биологических объектов заключаются в том, что в процессе их жизнедеятельности, в них возникают не скомпенсированные электрические заряды, которые в окружающем пространстве создают электрические поля. Эти поля, по отношению к биообъекту, называются внешними. Общеизвестно, что электрическое поле бесконечно в пространстве, хотя с увеличением расстояния от заряда, создающего поле, оно ослабляется. Проще всего оценить величину внешних электрических полей можно, измерив разность потенциалов (напряжение) между двумя точками тела человека, как правило, наиболее удобных для размещения электродов. Такие потенциалы называются биопотенциалами действия органов или тканей. Графическую запись этих биопотенциалов с помощью устройств регистрации называют электрограммой, сокращенно ЭГ. Отведение биопотенциалов подчиняется закономерностям отведения биопотенциалов в объемном проводнике. Если источник ЭДС (клетка) погружен в объемный проводник (проводник второго рода), каким является мозг или другие ткани организма, а отводящий электрод представляет собой точку, находящуюся в этом проводнике, то принцип отведения значительно отличается от отведения, когда электроды расположены на поверхности нервного или мышечного волокна. При отведении с поверхности, когда оба электрода располагаются непосредственно возле источника, разность потенциалов между электродами будет отсутствовать до тех пор, пока несущий отрицательный заряд участок возбуждения не окажется под одним из электродов. В объемном проводнике, электрическое поле существует в среде все время, пока находящаяся в нем клетка активна. Расположенные в этой среде электроды будут при любом положении регистрировать активность источника.
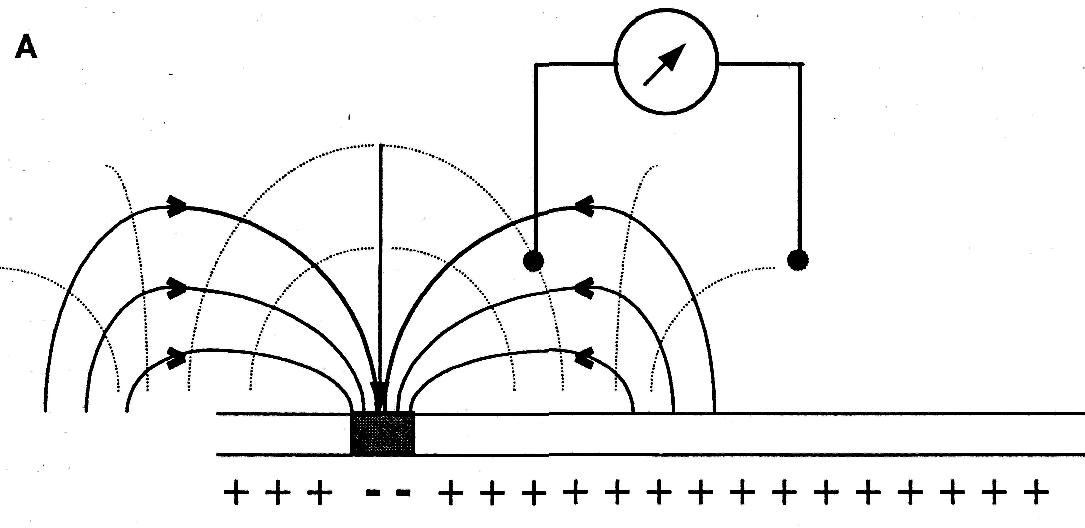
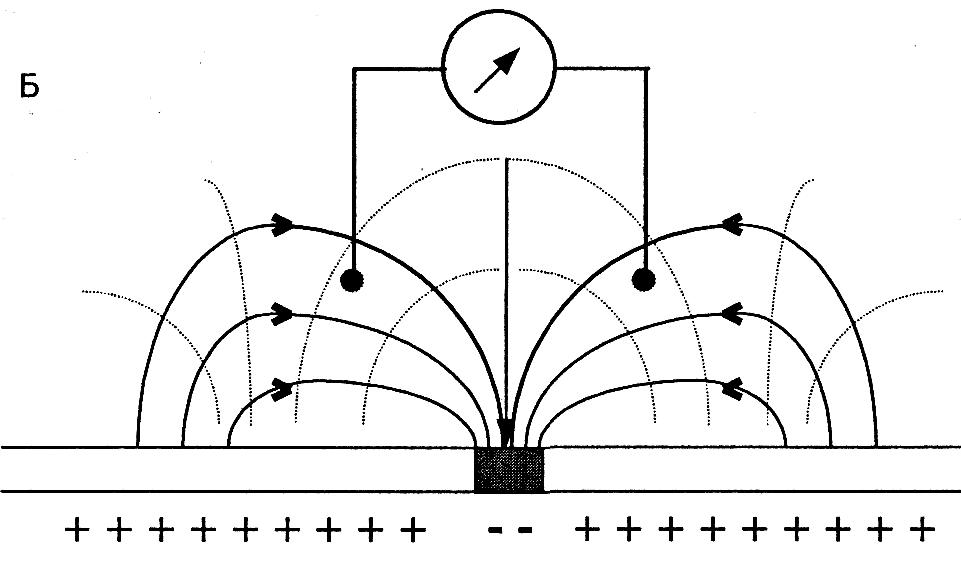
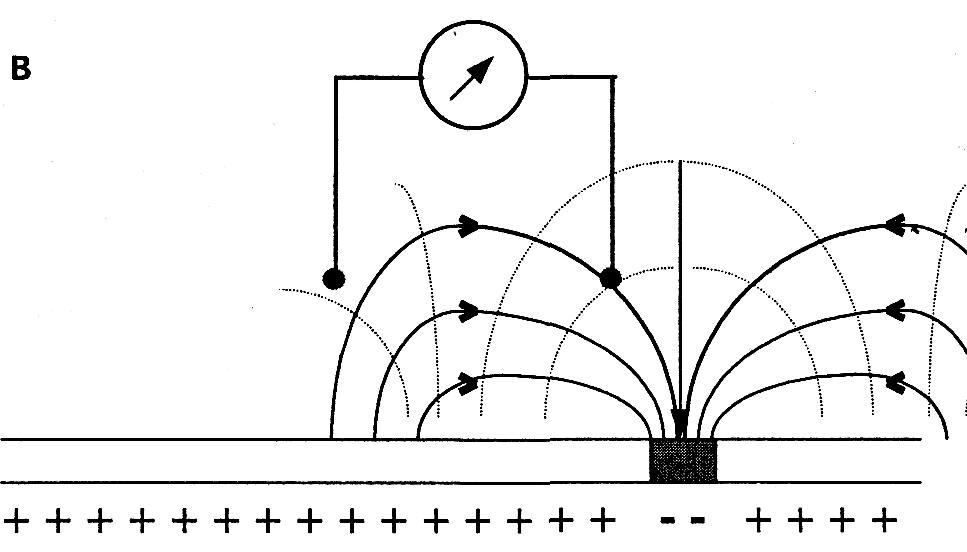
На рисунке показано распределение силовых линий и эквипотенциальных поверхностей в объемном проводнике вокруг волокна, при различных положениях области возбуждения. Меняющаяся разность потенциалов будет регистрироваться до тех пор, пока возбуждение проходит по волокну. При регистрации непосредственно с поверхности нервного волокна биопотенциалы возникают только при прохождении области возбуждения под электродом. Примером такого вида регистрации может быть ЭКГ, ЭЭГ, ЭМГ, ЭЭГ и другие. Все эти виды электрографии являются частными случаями записи биопотенциалов в объемном проводнике. Электрогенез этих электрограмм выходит за пределы данной лекции. Скажем только, что эти виды биопотенциалов являются суммарными т.е. складываются (синхронизируются, интегрируются) из биопотенциалов отдельных функциональных единиц (мышечных волокон сердца, мышечных волокон желудка, скелетной мускулатуры, нервных клеток головного мозга). Суммарный потенциал создает изменяющееся по напряженности электрическое поле, которое в объемном проводнике регистрируется в виде изменяющихся по величине биопотенциалов. Они имеют определенную форму, частоту, зависимую от функции органов. Отклонение параметров биопотенциалов от нормы при различных заболеваниях может служить объективными показателями для диагностики.
Читайте также: Эскизы картин из ткани
Поэтому возникает необходимость изучения электрограмм, и здесь ставятся две основные задачи:
1. Генез, т. е. формирование, механизм возникновения ЭГ.
2. Трактовка (сущность) ЭГ, т. е. отражение в них функции изучаемого органа в норме и при патологических изменениях.
Решение этих задач непосредственно на организме человека затруднено в связи с многообразием электрических процессов, происходящих в органах и тканях и их взаимовлиянием друг на друга. Поэтому генез и трактовку ЭГ изучают на физической модели, называемой эквивалентным электрическим генератором. Модель должна удовлетворять следующим условиям:
1.Расчетные потенциалы модели должны соответствовать в разных точках организма реально регистрируемым потенциалам.
2.При изменении параметров модели в ней должны происходить такие же изменения поля и его характеристик, как и в реальных ЭГ при нарушениях функционального состояния органа.
Модель эквивалентного электрического генератора представляет собой замкнутую электрическую цепь.
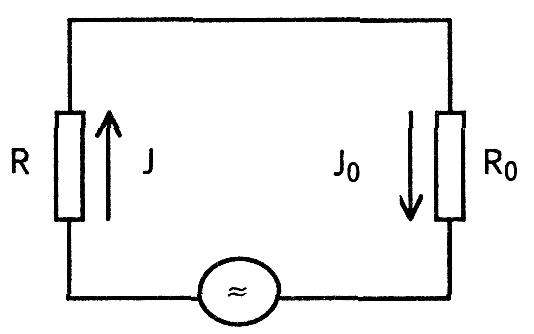
В эту цепь включены токовый генератор — источник ЭДС, имеющий собственное (внутреннее) сопротивление R. Сопротивление Ro (внешнее) соответствует сопротивлению тканей организма, в котором распространяется электрическое поле. Величина тока в замкнутой цепи одинакова через любой последовательный участок и будет определяться по закону Ома.
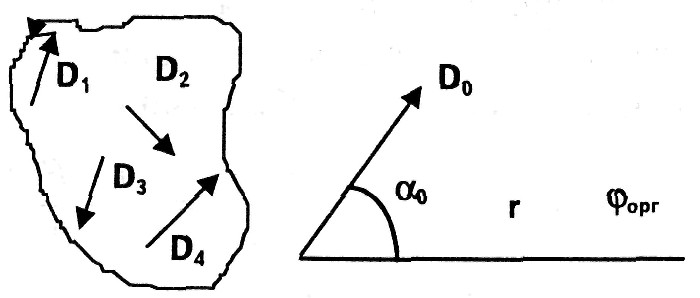
а т. к. реально R » R0, то J = J0 = ε / R, т.е. сила тока практически не зависит от сопротивления внешней среды. Элементарным токовым генератором в живой ткани является клетка. Модель токового генератора ткани, органа представляется в виде совокупности элементарных токовых генераторов (например, клеток), каждый из которых создает собственное электрическое поле. При этом имеет место суперпозиция (сложение) электрических полей отдельных микрогенераторов в общее поле органа. Пространственная структура и величина электрического поля, создаваемого токовым генератором, находящимся в проводящей среде, определяется положением полюсов генератора и свойствами среды. Для расчета потенциала в любой точке поля электрический генератор представляют в виде токового электрического диполя, т.е. системы положительного и отрицательного зарядов, определенным образом ориентированной в пространстве и находящейся в проводящей среде. Основной характеристикой диполя является электрический дипольный момент D.
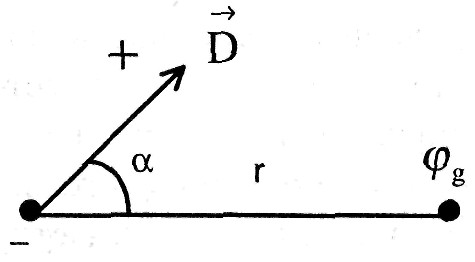
Дипольный момент — это вектор, определяющийся произведением тока диполя (равного суммарному току во внешней среде) на вектор расстояния между полюсами диполя(L).
Направление D от отрицательного полюса диполя к положительному.
Различают диполи точечные и конечные, точечные диполи имеют |L| → 0 . Рассматривают также электрические поля, создаваемые одним полюсом диполя, их называют униполями.
Рассмотрим значение потенциала, создаваемого точечным зарядом, элементарным диполем, органом тела человека. Будем считать, что окружающая их среда однородна в электрическом отношении.
1. Потенциал униполя в любой точке определяется по формуле:
где р — удельное сопротивление среды, J — ток, исходящий из данной точки униполя, r — расстояние от заряда до данной точки.

2. Потенциал поля, созданный элементарным диполем, определяется по формуле:
В этой формуле G есть сумма членов ряда, в котором каждое слагаемое пропорционально: L 2 /r 3 , L 3 /r 4 , L 4 /r 5 . Если r » L , то всеми членами ряда можно пренебречь и тогда
3. Электрическое поле, создаваемое каким-либо органом (например сердцем) равно геометрической сумме полей, создаваемых отдельными диполями, возникающими при работе этого органа. Потенциал поля в любой точке равен сумме потенциалов, создаваемых отдельными диполями
ri = const, т.к. размеры возбужденного участка незначительны по сравнению с расстоянием r. Можно доказать, что,
Приведенные выше количественные характеристики легли в основу одного из основных методов анализа возбудимости миокарда — многополярное отведение ЭКГ.
К пассивным электрическим свойствам биологических объектов относятся: сопротивление, электропроводимость, емкость, диэлектрическая проницаемость. В норме и патологии эти параметры меняются и поэтому могут быть использованы для изучения структуры и физико-химического состояния биологического вещества. Эти свойства проявляются, если к исследуемому участку ткани приложить напряжение небольшой величины.
Читайте также: Что такое ткань лен рогожка
При приложении постоянной разности потенциалов к тканям организма в них наблюдается два явления:
1. Постоянный электрический ток в проводящих тканях.
2. Различные виды поляризации в диэлектрических тканях. Величина тока в тканях определяется по закону Ома для участка цепи, однако для электролитов, а следовательно и биообъектов, закон имеет своеобразный вид:
В этой формуле U — приложенное к участку ткани напряжение, R — активное сопротивление этого участка, εn (t) – ЭДС поляризации, которая возникает в результате поляризационных явлений как на электродах, так и внутри ткани на полупроницаемых и непроницаемых для ионов перегородках. ЭДС поляризации со временем возрастает, а ток в тканях уменьшается и при длительном воздействии становится равным нулю
.
В диэлектриках заряды связаны, однако они перемещаются при наложении внешнего электрического поля внутри микроструктуры: атома, молекулы, клетки или в пределах границы проводящей и непроводящей среды. Для каждого вида поляризации приводится значение времени релаксации τ.
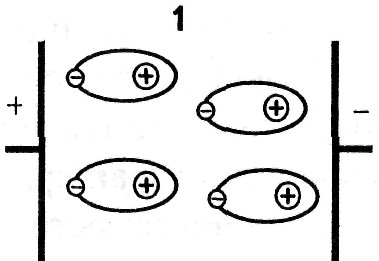
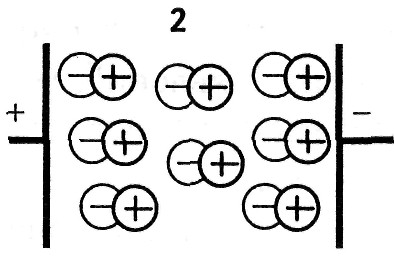
Время релаксации — это время, в течение которого поляризация увеличивается от нуля до максимума, с момента приложения внешнего напряжения.
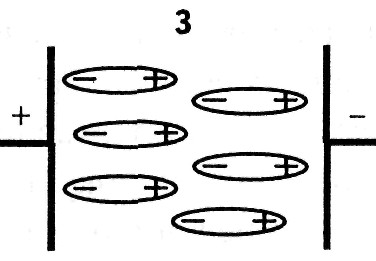
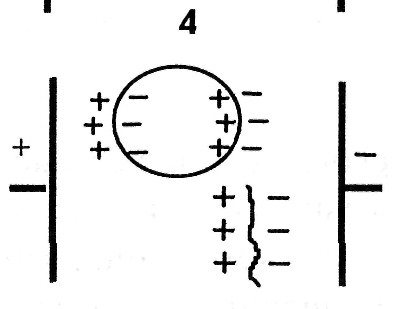
1. При электронной поляризации под воздействием внешнего электрического поля происходит деформация электронных орбиталей атомов, ориентированных вдоль поля. Время релаксации = (10 -16 — 10 -14 ) с.
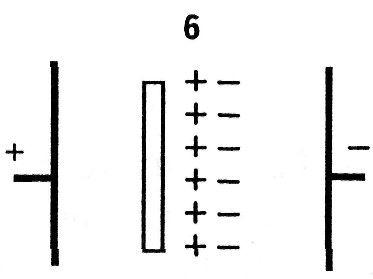
2. При ионной поляризации происходит смещение ионов в кристаллической решетке вдоль направления электрического поля, = (10 -8 — 10 -3 )с.
3. Дипольно-ориентационная поляризация происходит в структурах, в которых уже имеются полярные молекулы — диполи, ориентированные хаотично. Под действием электрического поля они выстраиваются вдоль поля, = (10 -13 — 10 -7 ) с.
4. При микроструктурной поляризации происходит перераспределение ионов в результате действия электрического поля на различных полупроницаемых и непроницаемых для ионов перегородках, например: на цитоплазматических мембранах, мембранах клеточных органоидов, некоторых разделительных тканевых оболочках. В результате такого перераспределения возникает структура, подобная гигантской поляризованной молекуле, = (10 -8 — 10 -3 ) с.
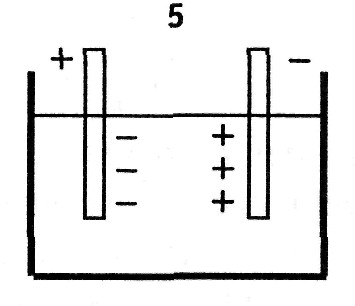
5. Электролитическая или электрохимическая поляризация возникает между электродами, опущенными в электролит. Ионы, подходящие к электродам, не полностью успевают нейтрализоваться по причине вторичных реакций на электродах и неодинаковой подвижности ионов. В результате, вокруг каждого электрода возникает «облако» зарядов противоположного знака, что ведет к образованию поля, направленного противоположно внешнему и постепенному уменьшению тока, проходящего через электролит, = (10 -3 — 10 2 ) с.
6. Поверхностная поляризация возникает на образованиях, имеющих двойной электрический слой. Ионы дисперсионной части двойного электрического слоя связаны с атомами поверхности и не являются свободными. Диффузионный слой образуется за счет притяжения ионами дисперсионного слоя. При приложении внешнего поля происходит частичное смещение ионов обеих слоев, образуются так называемые наведенные диполи, = (10 -3 — 1) с.
Все рассмотренные явления поляризации в той или иной степени присущи биологическим объектам. При приложении внешнего поля в тканях индуцируется противоположно направленное поле за счет поляризационных явлений, которое уменьшает внешнее поле и обуславливает высокое удельное сопротивление тканей постоянному току. Количественно явление поляризации характеризуется величиной относительной диэлектрической проницаемости.
где Ео — напряженность внешнего электрического поля в вакууме, Е — напряженность поля в среде, оно равно разности Ео и Еn, где Еn — напряженность поля, создаваемая наведенными диполями при поляризации. Относительную диэлектрическую проницаемость можно определить также соотношением емкостей
где Со — емкость электродов в вакууме, С — емкость электродов в среде.
При приложении к биологическому объекту переменного (как правило, синусоидального) напряжения, в нем также возникают электрический ток и поляризационные явления. Электрическую модель биологического объекта для переменного тока можно представить в виде двух сопротивлений:
— индуктивное сопротивление равно нулю.
Эти сопротивления в самой ткани могут быть соединены как последовательно так и параллельно.
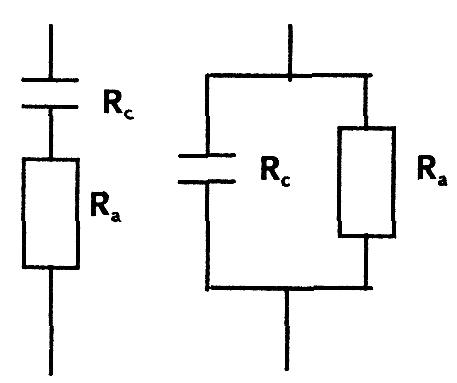
Общее сопротивление ткани в цепи переменного тока называется импедансом и обозначается Z. Импеданс определяется по формуле:
— при последовательном соединении
— при параллельном соединении:
Импеданс ткани изменяется с частотой. Это явление называют дисперсией. Величина импеданса определяется сопротивлением самой ткани, а также зависит от соотношения частоты или перио-
да приложенного напряжения и времени релаксации:
1) если Т/4 > τ (Т/4 — время возрастания приложенного напряжения от 0 до max, τ — время релаксации), проводимость объекта и диэлектрическая проницаемость с частотой не меняется,
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом