
ХЕМОРЕЦЕПТОРЫ (средневековое арабское al-kimiya; греческий chymeia и chemeia искусство сплавления металлов + рецепторы; синоним хемоцепторы) — специальные чувствительные образования, воспринимающие воздействия химических раздражителей и преобразующие их в нервные сигналы.
Процесс взаимодействия химического стимула с рецепторной мембраной хемочувствительной клетки, приводящей к возбуждению нейрона, называется хеморецепцией. Механизм этого процесса мало изучен. Считают, что за. хеморецепцию ответственны специальные макромолекулы, входящие в состав мембраны хеморецепторов. Так, из ткани языка млекопитающих, содержащей вкусовые рецепторы, выделены мембранные фракции, включающие макромолекулы, способные взаимодействовать со сладкими и горькими веществами.
Среди хеморецепторов различают экстероцепторы, воспринимающие химические раздражения извне (вкусовые и обонятельные рецепторы), и интероцепторы, воспринимающие изменения химического состава крови, лимфы и других тканей. Возбуждение вкусовых рецепторов вызывает ощущение вкуса (см.), а обонятельных рецепторов — запаха (см.). Возбуждение хеморецепторов внутренних органов не всегда сопровождается возникновением четких образных восприятий. Возникающие при этом субъективные ощущения были названы И. М. Сеченовым «системными чувствами» — голод, жажда и т. д. (см. Интероцепция).
Морфологически хеморецепторы весьма разнообразны. Из хеморецепторов внутренних органов наиболее обстоятельно изучены артериальные хеморецепторы, расположенные в главных артериальных магистралях,— в сонной артерии и в дуге аорты. Хеморецепторы внутренней сонной артерии сосредоточены в каротидном гломусе (см. Параганглии). Хеморецепторы дуги аорты представляют собой разрозненные скопления клеток. Каротидный гломус имеет сложную капиллярную структуру с многочисленными артериовенозными анастомозами. Такие анастомозы считают отличительной особенностью тканевых хеморецепторов. Между капиллярами располагаются клетки двух типов: клетки I типа заполнены электронно-плотными везикулами, содержащими биогенные амины (см.); окружающие их своими цитоплазматическими отростками клетки II типа не содержат биогенных аминов. На клетках I типа образуют синаптические контакты окончания волокон каротидного синусного нерва являющегося ветвью языкоглоточного нерва (см.), который содержит как афферентные, так и эфферентные волокна. Собственно хеморецепторами в каротидном гломусе считаются клетки I типа и свободные хемочувствительные нервные окончания.
Артериальные хеморецепторы реагируют на изменение концентрации в крови кислорода, углекислоты и водородных ионов. Гипоксия (см.) гиперкапния (см.), сопровождающиеся изменениями хим. состава крови (см. Кровь), приводят к возникновению сердечно-сосудистых и дыхательных рефлексов, направленных на нормализацию состава крови и поддержание гомеостаза (см.). Поэтому основной функцией этих хеморецепторов является участие в регуляции легочной вентиляции (см.), обеспечивающей оптимальное соотношение кислорода и углекислоты в крови. Каротидные хеморецепторы играют большую роль в регуляции дыхания (см.), чем аортальные хеморецепторы, участвующие преимущественно в регуляции деятельности сердечнососудистой системы (см.).
Механизм стимуляции артериальных хеморецепторов связан с особенностями кровоснабжения органа. Так, импульсация в синусном нерве, вызванная раздражением хеморецепторов каротидного гломуса, зависит не столько от парциального напряжения кислорода и углекислоты в артериальной крови, сколько от концентрации кислорода и водородных ионов в ткани самого каротидного гломуса. Возбуждение артериальных хеморецепторов может происходить также под влиянием некоторых неорганических и органических веществ. На этом основано, например применение лобелина (см.) для стимуляции дыхания. Физиологическое значение хеморецепторов, локализованных в тканях и органах (печени, желудке, кишечнике и др.), заключается в восприятии органами и тканями концентрации питательных веществ, гормонов, осмотического давления крови и передаче сигнала об их изменениях в центральной нервной системе.
Нарушение хеморецепции может быть вызвано патологическими процессами в органах и тканях, где расположены хеморецепторы (например, при глосситах, глоссалгии), а также патологическими процессами в других физиологических системах организма. Так, например, вторичное нарушение функций вкусовых хеморецепторов наблюдается при хронических заболеваниях желудочно-кишечного тракта (гастриты, колиты и др.).
Читайте также: Как убрать воск с ткани батик
Экспериментальные методы изучения хеморецепторов внутренних органов основаны на регистрации рефлекторных реакций, возникающих в ответ на воздействие тех или иных химических веществ на органы и ткани или при их перфузии через гуморально изолированные органы (см. Перфузия). Для исследования хеморецепторов используют также электрофизиологическую регистрацию импульсной активности с афферентных волокон, отходящих от органов и тканей (см. Электрофизиология).
Библиогр.: Лебедева В. А. Механизмы хеморецепции, М.— Л., 1965; Самойлов В. О. Некоторые проблемы каротидной хеморецепции, в кн.: Сенсорные системы, Морфофизиологические и поведенческие аспекты, под ред. Г. В. Гершуни, с. 76, Л., 1977; Физиология кровообращения, под ред. Е. Б. Бабского и др., Л., 1980; Handbook of sensory physiology, ed. by E. Neil, v. 3, pt 1, В. a. o., 1972; The peripheral arterial chemorecep-tors, ed. by M. J. Purves, Cambridge, 1975.
ПЕРИФЕРИЧЕСКИЕ РЕЦЕПТОРЫ Периферические хеморецепторы
К периферическим хеморецепторам относятся ка-ротидные тельца (расположенные в области бифуркации общей сонной артерии) pi аортальные тельца (расположенные в области дуги аорты). Ka-ротидные тельца — это главные периферические хеморецепторы в организме человека, они реагируют на изменения PaO2, PaCO2, рН и артериального перфузнойного давления. Они связаны с дыхательными центрами через языкоглоточный нерв и обеспечивают рефлекторное увеличение альвеолярной вентиляции при снижении PaO2, артериальной перфузии, а также при повышении [H + ] или PaCO2. Периферические хеморецепторы также чувствительны к цианидам, доксапраму и высоким дозам никотина. В отличие от центральных хемо-рецепторов, которые воспринимают главным образом изменения PaCO2 (в действительности [H + ]), каротидные тельца наиболее чувствительны к PaO2 (рис. 22-26). Заметим, что активность рецепторов существенно не меняется, пока PaO2 не упадет ниже 50 мм рт. ст. Считается, что клетки каротидных телец (гломусные клетки) представляют собой дофаминергические нейроны. Анти-дофаминергические препараты (фенотиазины), большинство анестетиков, а также хирургическое вмешательство на обеих сонных артериях приводят к исчезновению периферического рефлекторного дыхательного ответа на гипоксемию.
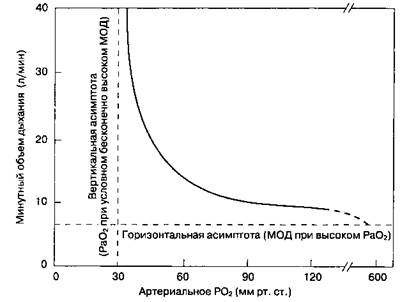
Рис. 22-26.Зависимость между PaO2 и минутным объемом дыхания (МОД) в покое и при нормальном PaCO2. (С разрешения. Из: Nunn J. F. Applied Respiratory Physiology, 3rd ed. Butterworths, 1987.)
Легочные рецепторы
Импульсы от этих рецепторов поступают в центральную нервную систему по блуждающему нерву. Рецепторы растяжения располагаются в гладких мышцах дыхательных путей; ohpi ответственны за прекращение вдоха, когда легкие перераздуты (инфляционный рефлекс Геринга-Брей-ера),и укорочение выдоха, когда легкие спадаются (дефляционный рефлекс).В обычных условиях у человека рецепторы растяжения играют незначительную роль. Двусторонняя блокада блуждающих нервов практически не сказывается на нормальном дыхании.
Ирритантные рецепторы, расположенные в слизистой оболочке трахеи и бронхов, реагируют на раздражающие газы, сигаретный дым, пыль и холодный воздух; pix актршация вызывает рефлекторное учащение дыхания, бронхоконстрикцию и кашель. J-рецепторы (юкстакапиллярные) расположены в интерстициальном пространстве внутри альвеолярной стенки; эти рецепторы играют роль в возникновении одышки в ответ на увеличение объема ирггерстирщального пространства и на воздействие различных химических медиаторов, выде-ляющихся при повреждении легочной ткани.
Другие рецепторы
К таковым относятся различные мышечные и суставные рецепторы дыхательной мускулатуры и грудной клетки. Импульсы из этих источников,
возможно, важны во время физической нагрузки и прр! патологических состояниях, сопровождающихся уменьшением растяжимости легких рыи грудной клетки.
Читайте также: Белки опорных тканей это
Периферические хеморецепторы располагаются в гломусной ткани каротидных телец
или Пневмапсихосоматология человека
Русско-англо-русская энциклопедия, 18-е изд., 2015
(Греч.: καρόω — погружать в глубокий сон, лишать чувств, оглушать; 1940).
Каротидное тельце — это парный орган артериальной хеморецепции, расположенный в области каротидного синуса, в задней стенке общей сонной артерии в месте ее разделения на внутреннюю и наружную сонную артерию.
Каротидное тельце представляет собой структуру красновато-коричневого цвета овальной формы диаметром приблизительно 5 мм, окруженную фиброзной капсулой.
Xеморецепторы каротидных телец подобны хеморецепторам аортальных телец. Они реагируют на снижение в крови уровня концентрации кислорода или уровня pH, а также косвенно (за счет изменения pH) на изменение уровня концентрации в крови двуокиси углерода. Информация о химических характеристиках крови передается по афферентным волокнам языкоглоточного нерва в центральную нервную систему и используется регуляторами вегетативных систем для управления дыханием, кровообращением и другими функциями организма.
Схема. Долька каротидного тельца (крыса).
Модификация: McDonald DM, Mitchell RA (1975) The innervation of glomus cells, ganglion cells and blood vessels in the rat carotid body: a quantitative ultrastructural analysis. J. Neurocytol. 4:177-230.
Каротидные тельца состоят из множества долек — «клубочков» (см. схему слева). Клубочки окружают кровеносные капилляры каротидного тельца. Клубочек состоит из нескольких гломерулярных клеток (клетки типа I) и, окружающих гломерулярные клетки, поддерживающих клеток (клетки типа II). Клубочки обильно снабжаются кровью. О высокой их активности свидетельствует интенсивность метаболизма (высокий уровень потребления кислорода на единицу массы).
Микрогемациркуляторное русло клубочков представляет собой сложную сеть мелких и крупных капилляров и артериовенозных шунтов. Артериолы, дающие начало микрогемациркуляторному руслу, иннервируются симпатическими эфферентными нервными волокнами и парасимпатическими постганглионарными эфферентными волокнами.
Полагают, что именно гломерулярные клетки являются сенсорными рецепторами. Они преобразуют информацию о химических характеристиках протекающей через клубочки крови в вероятностную последовательность нервных импульсов. Гломерулярные клетки клубочков имеют яйцевидную форму. Их цитоплазма содержит крупное ядро и множественные гранулы (пузырьки) с плотной сердцевиной. Гломерулярные клетки могут иметь цитоплазматические отростки различной длины и образовывать между собой сложные соединения.
К клубочкам подходят афферентные (чувствительные) волокна языкоглоточных нервов (веточка языкоглоточного нерва — синусный нерв, или нерв Геринга). Терминали этих волокон образуют синапсы на некоторых гломерулярных клетках. Одно нервное волокно может иннервировать 10 ÷ 20 гломерулярных клеток. Многие гломерулярные клетки не иннервируются. Некоторые гломерулярные клетки могут иннервироваться эфферентными симпатическими волокнами, идущими от симпатического сплетения сонной артерии.
Характер отношений гломерулярных клеток друг с другом и с терминалями нервных волокон приводят к ряду предположений о назначении и функциях гломерулярных клеток. Одни гломерулярные клетки могут выполнять функцию сенсорных хеморецепторов — структур, воспринимающих информацию о химическом составе крови, протекающей по микрогемациркуляторному руслу клубочков. Другие гломерулярные клетки могут выполнять функцию модуляторов чувствительности хеморецепторов. И, наконец, гломерулярные клетки могут выполнять функцию модуляторов возбудимости терминалей афферентных волокон, передающих информацию в центральную нервную систему.
Периферические хеморецепторы располагаются в гломусной ткани каротидных телец
Хеморецепторный контроль дыхания осуществляется при участии центральных и периферических хеморецепторов. Центральные (медуллярные) хеморецепторы расположены непосредственно в в ростральных отделах вентральной дыхательной группы, в структурах голубого пятна (locus coeruleus), в ретикулярных ядрах шва ствола мозга и реагируют на водородные ионы в окружающей их межклеточной жидкости мозга (рис. 10.23). Центральные хеморецепторы представляют собой нейроны, которые в определенной степени являются рецепторами углекислого газа, поскольку величина рН обусловлена парциальным давлением С02, согласно уравнению Гендерсона—Гасельбаха, а также тем, что концентрация ионов водорода в межклеточной жидкости мозга зависит от парциального давления углекислого газа в артериальной крови.
Читайте также: Ткани мира в ясенево
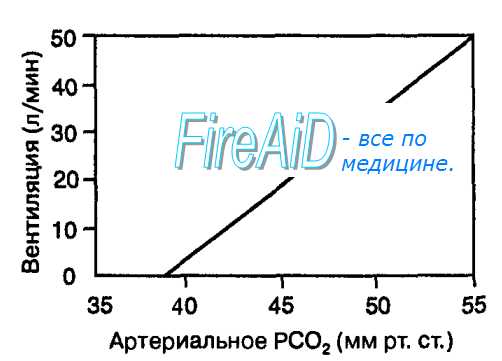
Рис. 10.23. Зависимость вентиляции легких от степени стимуляции центральных хеморецепторов изменениями [Н+]/РС02 в артериальной крови. Увеличение парциального давления С02 в артериальной крови выше порога (РС02 = 40 мм рт. ст.) линейно увеличивает объем вентиляции легких.
Увеличение вентиляции легких при стимуляции центральных хеморецепторов ионами водорода называется центральным хеморефлексом, который оказывает выраженное влияние на дыхание. Так, в ответ на уменьшение рН внеклеточной жидкости мозга в области локализации рецепторов на 0,01 легочная вентиляция возрастает в среднем на 4,0 л/мин. Однако центральные хеморецепторы медленно реагируют на изменения С02 в артериальной крови, что обусловлено их локализацией в ткани мозга. У человека центральные хеморецепторы стимулируют линейное увеличение вентиляции легких при увеличении С02 в артериальной крови выше порог, равного 40 мм рт. ст.
Периферические (артериальные) хеморецепторы расположены в каротидных тельцах в области бифуркации общих сонных артерий и в аортальных тельцах в области дуги аорты. Периферические хеморецепторы реагируют как на изменение концентрации водородных ионов, так и парциального давления кислорода в артериальной крови. Рецепторы чувствительны к анаэробным метаболитам, которые образуются в ткани каротидных телец при недостатке кислорода. Недостаток кислорода в тканях каротидных телец может возникнуть, например, при гиповентиляции, ведущей к гипоксии, а также при гипо-тензии, вызывающей снижение кровотока в сосудах каротидных телец. При гипоксии (низкое парциальное давление кислорода) периферические хеморецепторы активируются под влиянием увеличения концентрации в артериальной крови, прежде всего, ионов водорода и РС02.
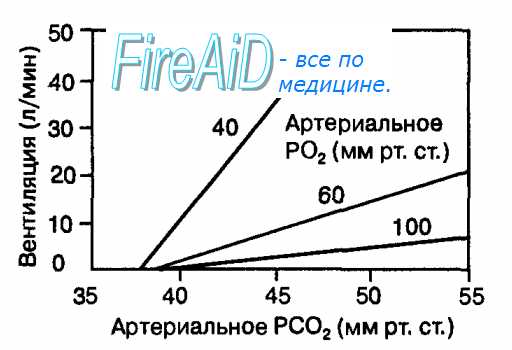
Рис. 10.24. Зависимость вентиляции легких от степени стимуляции периферических хеморецепторов гипоксическим стимулом. При стимуляции периферических хеморецепторов гипоксией имеет место мультипликативное взаимодействие парциального давления С02 в артериальной крови и гипоксии, в результате которого происходит максимальное увеличение вентиляции легких. Напротив, при высоком Парциальном давлении кислорода в артериальной крови периферические хеморецепторы слабо реагируют на увеличение РС02. Если в артериальной крови парциальное давление С02 становится ниже порога (40 мм рт.ст.), то периферические хеморецепторы также слабо реагируют на гипоксию.
Действие на периферические хеморецепторы этих раздражителей усиливается по мере снижения в крови Р02 (мультипликативное взаимодействие). Гипоксия увеличивает чувствительность периферических хеморецепторов к [Н+] и С02. Это состояние называется асфиксией и возникает при прекращении вентиляции легких. Поэтому периферические хеморецепторы называются часто рецепторами асфиксии. Импульсы от периферических хеморецепторов по волокнам синокаротидного нерва (нерв Геринга — часть языкоглоточного нерва) и аортальной ветви блуждающего нерва достигают чувствительных нейронов ядра одиночного тракта продолговатого мозга, а затем переключаются на нейроны дыхательного центра. Возбуждение последнего вызывают прирост вентиляции легких. Вентиляция легких увеличивается линейно в соответствии с величиной [Н+] и РС02 выше порога (40 мм рт. ст.) в артериальной крови, протекающей через каротидные и аортальные тельца (рис. 10.24). Наклон кривой на рисунке, который отражает чувствительность периферических хеморецепторов к [Н+] и РС02, варьирует в зависимости от степени гипоксии.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом