
Удивительно, как в поисках источников электрического тока пересекаются судьбы исследователей. Один ученый случайно открыл «животное электричество», изучая анатомию лягушки, а другой нашел способ получать электрический ток с помощью химической реакции.

Луиджи Гальвани едва ли был физиком. Он последовал за своим отцом в медицину и, нарабатывая навыки хирурга, много занимался анатомией. И прежде чем начать работать с пациентами, тренировался на мертвых животных. Луиджи даже поработал на полной ставке анатома в Болонском университете.
После девяти лет академических исследований Гальвани совершил открытие, в котором есть немалая доля случайности. Он повесил пару лягушачьих лапок на проволочную сетку для просушки. Сетка была из железа, а крючки для подвеса из меди. Неожиданно свежие лягушачьи лапки начали подергиваться. По некоторым отчетам, ученый даже заметил искру!
Гальвани обнаружил, что может повторить этот эффект, пользуясь заряженной лейденской банкой, когда попытался стимулировать «живую» мускулатуру (по крайней мере недавно погибшего животного) электрическим разрядом. Этот эффект Гальвани назвал «животным электричеством».
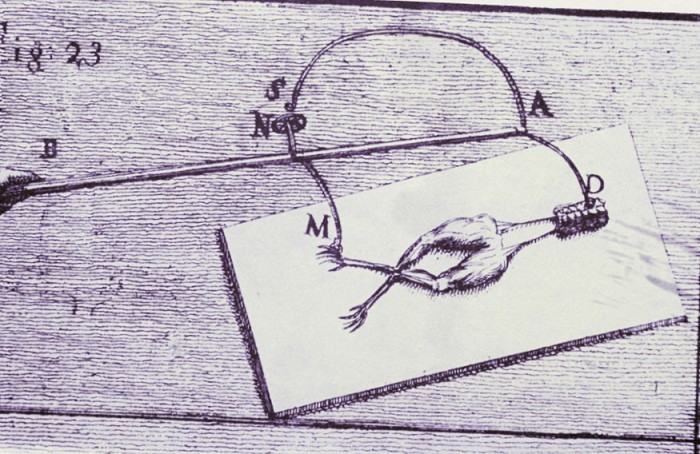
Исследования Гальвани этого интересного явления (когда лягушачьи лапки дергались на сетке) нашли в физической науке необычайно широкий отклик. Впоследствии Луиджи воспроизвел ситуацию, возникшую в его лаборатории, взяв металлическую дугу из двух металлов — меди и железа — и прикоснувшись скруткой к обнаженному позвоночнику лягушки (именно там располагаются нервы, которые контролируют мышцы ноги) и к кончику лапки.
В своем опыте Гальвани создал электрическую цепь, по которой «животное электричество» прошло через мускулатуру лягушки, заставляя ее сокращаться. Но где рождалось это электричество? Гальвани предполагал, что он открыл некоторую форму «витальной силы», характерной для живых существ, однако 30 лет спустя другой ученый показал, как получить тот же эффект без животного.
Электрическая батарея
Металлическая дуга Гальвани вырабатывала ток только в контакте со свежим мясом, которое, как предполагал Гальвани, дает флюиды. Итальянец Алессандро Вольта заменил мясо деревянными опилками, размоченными в соленой воде. Он понял, что самым главным здесь являются два металла, которые реагируют друг с другом, за счет чего электрический заряд переходит из одного металла в другой.

Этот эффект Вольта попытался усилить, сделав в своей конструкции много биметаллических элементов, буквально входящих друг в друга. Его первый «вольтов столб» представлял собой серебряные монеты, переложенные цинковыми дисками и разделенные сырыми древесными опилками.
Соединение проводом верхней монеты с нижним диском приводило к тому, что по цепи тек электрический ток. Таким образом, «животное электричество» Гальвани оказалось тем же самым явлением, что и «тепловое электричество» Вольты (он сам так назвал свое открытие, поскольку столб разогревался), и, чтобы разобраться в этом, физикам придется открыть еще немало законов природы.
Батарейка
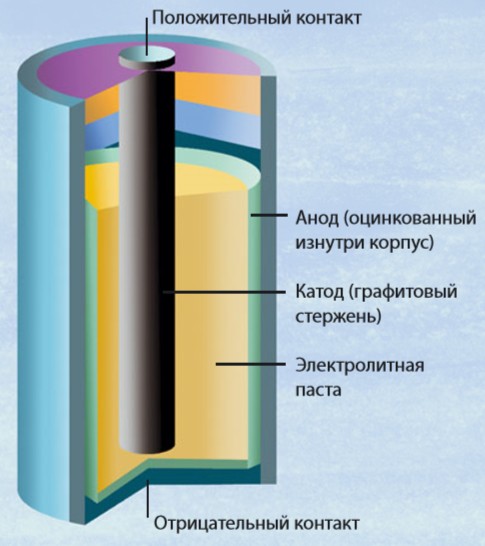
Современная батарейка работает по тем же самым принципам, что и «вольтов столб». Два вещества (катод и анод) подобраны так, чтобы химически взаимодействовать друг с другом. В процессе этого взаимодействия электроны переходят от анода к катоду. Устройство батарейки таково, что две взаимодействующие части не имеют контакта друг с другом, поэтому электроны между ними движутся через жидкость, называемую электролитом. Так создается электрический ток.
Франкенштейн

Племянник и последователь Гальвани, Джованни Альдини превратил опыты с животным электричеством в шоу «электрические пляски». Он ездил с турами по Европе, покупая тела недавно казненных и заставляя их трястись под действием электричества под шумные аплодисменты публики. Как говорят, автор романа «Франкенштейн, или Современный
Прометей» 18-летняя Мэри Шелли посвятила этот роман монстру, оживленному электричеством, под впечатлением жутких гальванических опытов Альдини.
Первым исследователем электрической активности живых тканей является л гальвани
В учебнике в соответствии с программой представлены все разделы физиологии человека. Наиболее полно изложены главы «Вегетативная нервная система», «Железы внутренней секреции», «Центральная нервная система», «Анализаторы». В главе «Физиология дыхания» дается новая полная классификация гипоксии, созданная одним из авторов. В учебнике впервые излагаются основы физиологических функций организма в связи с данными по фармакологической коррекции их нарушений. Для студентов медицинских и биологических факультетов и медицинских институтов, обучающихся по специальностям «Лечебное дело», «Стоматология» и «Фармация».
Читайте также: Обшить круглый стул тканью своими руками
Николай Александрович Агаджанян
Глава 1. История физиологии. Методы физиологических исследований
Глава 2. Физиология возбудимых тканей
Биоэлектрические явления в возбудимых тканях. Природа возбуждения
Изменения мембранного потенциала. Пороговые и подпороговые раздражители
Изменения возбудимости при возбуждении
Законы раздражения возбудимых тканей
Физиология нервов и нервных волокон
Механизм мышечного сокращения
Фармакологические влияния на возбудимые ткани
Глава 3. Физиология центральной нервной системы
Организация нервной системы
Общие закономерности деятельности центральной нервной системы
Рефлекторный принцип регуляции
Торможение в центральной нервной системе и его виды
Классификация видов торможения
Принципы координационной деятельности центральной нервной системы
Частная физиология центральной нервной системы
Собственные функции спинного мозга
Проводниковая функция спинного мозга
Собственные функции продолговатого мозга
Вегетативные функции продолговатого мозга
Проводниковые функции продолговатого мозга
Собственные функции варолиева моста
Проводниковая функция варолиева моста
Проводниковая функция гипоталамуса
Собственные функции гипоталамуса
Функции лимбической системы
Локализация функций в коре больших полушарий
Электрическая активность коры головного мозга
Функции гематоэнцефалического барьера
Факторы, повышающие проницаемость гематоэнцефалического барьера
Особенности морфологического строения гематоэнцефалического барьера
Фармакологические препараты, регулирующие функцию центральной нервной системы
Глава 4. Вегетативная (автономная) нервная система
Различия между вегетативной и соматической нервными системами
Структура и функции вегетативной нервной системы
Симпатический отдел вегетативной нервной системы
Парасимпатический отдел вегетативной нервной системы
Внутриорганный отдел (энтеральный, метасимпатический)
Медиаторы вегетативной нервной системы
Вегетативные (автономные) рефлексы
Центры регуляции вегетативных функций
Средства, влияющие на синаптическую передачу
Глава 5. Железы внутренней секреции
Общая физиология желез внутренней секреции
Механизмы действия гормонов.
Регуляция функций желез внутренней секреции
Частная физиология желез внутренней секреции
Гормоны передней доли гипофиза
Гормоны задней доли гипофиза
Околощитовидные (паращитовидные) железы
Гормоны коры надпочечников
Гормоны мозгового слоя надпочечников
Мужские половые гормоны (андрогены)
Овариально-менструальный (менструальный) цикл
Гормональные средства, используемые в фармакологические целях
Объем и физико-химические свойства крови
Кислотно-основное состояние крови (КОС).
Гемоглобин и его соединения
Скорость оседания эритроцитов (СОЭ)
Фармакологическая коррекция нарушений гемопоэза и гемостаза
Средства, влияющие на гемопоэз
Средства, влияющие на гемостаз
Глава 7. Крово- и лимфообращение
Электрическая активность клеток миокарда и проводящей системы сердца
Проводимость и сократимость
Классификация сосудов. Основы гемодинамики
Транссосудистый обмен веществ
Нейрогуморальная регуляция кровообращения
Регуляция деятельности сердца
Внутрисердечные механизмы регуляции
Характер влияний блуждающих и симпатических нервов на работу сердца
Гуморальная регуляция деятельности сердца
Местные регуляторные механизмы
Центральные механизмы регуляции
Гуморальная регуляция сосудистого тонуса
Рефлекторная регуляция деятельности сердца и сосудистого тонуса
Методы исследования сердечно-сосудистой системы
Регуляция коронарного кровотока
Функции лимфатической системы
Нервная регуляция лимфообразования
Гуморальная регуляция лимфотока и лимфообразования
Фармакологическая коррекция нарушений некоторых физиологических показателей системы кровообращения
Средства, влияющие на возбудимость, проводимость сердечной мышцы и ритм сердечных сокращений
Средства, влияющие на сократимость сердечной мышцы
Средства, улучшающие коронарный кровоток и метаболизм миокарда
Средства, нормализующие кровяное давление
Средства, влияющие на метаболизм сосудистой стенки и ее проницаемость
Глава 8. Физиология дыхания
Состав и свойства дыхательных сред
Внутриплевральное и внутрилегочное давление
Вентиляция легких и легочные объемы
Газообмен и транспорт газов
Регуляция внешнего дыхания
Локализация и функциональные свойства дыхательных нейронов
Рефлекторная регуляция дыхания
Рефлексы с проприорецепторов дыхательных мышц
Гуморальная регуляция дыхания
Дыхание в измененных условиях
Дыхание при высоком атмосферном давлении
Патологические типы дыхания
Негазообменные функции воздухоносных путей и легких
Фармакологическая коррекция патологии органов дыхания
Функции желудочно-кишечного тракта
Общие принципы регуляции процессов пищеварения
Первым исследователем электрической активности живых тканей является л гальвани
Тема: Физиология возбудимых тканей
I . История открытия животного электричества.
Электрические явления в животных организмах известны давно. Еще в 1776 г. они были описаны у электрического ската. Началом же экспериментального изучения электрических явлений в животных тканях следует считать опыты итальянского врача Луиджи Гальвани (1791). В опытах он использовал препараты задних лапок лягушки, соединенных с позвоночником. C уществует легенда, что Л. Гальвани готовил лягушачьи лапки своей больной жене на обед, и идея пришла ему в голову, когда он их просто сгибал туда-сюда. Подвешивая эти препараты на медном крючке к железным перилам балкона, он обратил внимание, что, когда конечности лягушки раскачивались ветром, их мышцы сокращались при каждом прикосновении к перилам. На основании этого Гальвани пришел к выводу, что подергивания лапок были вызваны «животным электричеством», зарождающимся в спинном мозге лягушки и передаваемым по металлическим проводникам (крючку и перилам балкона) к мышцам конечностей.
Читайте также: Цветы из ткани своими руками схемы шаблоны выкройки
В 1791г. Л. Гальвани опубликовал свой труд «Трактат о силах электричества при мышечном движении». Однако, против положения Л. Гальвани о «животном электричестве» выступил физик Александр Вольта. В 1792 г. Вольта повторил опыты Гальвани и установил, что описанные Гальвани явления нельзя считать «животным электричеством». В опыте Гальвани источником тока служил не спинной мозг лягушки, а цепь, образованная из разнородных металлов, — меди и железа.
Вольта был прав. Первый опыт Гальвани не доказывал наличия «животного электричества», но эти исследования привлекли внимание ученых к изучению электрических явлений в живых образованиях.
В ответ на возражение Вольта Гальвани произвел второй опыт, уже без участия металлов. Конец седалищного нерва он набрасывал, стеклянным крючком на мышцу конечности лягушки; при этом также наблюдалось сокращение этой мышцы.
Племянник Гальвани Д. Альдини был пылким сторонником своего дяди и считал, что Вольта заблуждается. Он в своих затеях зашел так далеко, что во время наполеоновских войн стал ставить опыты на головах только что гильотированных людей. Как только голова отлетала, Альдини тут же ее подхватывал и прикладывал ток к ушам и губам, и на отрубленной голове появлялись различные гримасы.
В 1841 г свои исследования в области физиологии начал знаменитый немецкий физиолог Дюбуа Реймон, который считается «отцом» электрофизиологии. Он ввел в электрофизиологию термины, которыми пользуются до сих пор, например, ток покоя. Д. Реймон предполагал, что мышца и нерв ориентированы так, что их продольные участки заряжены положительно, а поперечные – отрицательно. При всяком раздражении происходит поворот этих молекул на 90 0 и тем самым происходит распространение тока.
Впервые В. .Ю. Чаговец в 1896 году высказал гипотезу об ионном механизме электрических потенциалов в живых клетках и сделал попытку объяснить их с позиции теории электролитической диссоциации С. Аррениуса. В 1902 году Ю. Бернштейном была разработана мембранно–ионная теория, согласно которой потенциал покоя нервных и мышечных волокон определяется избирательной проницаемостью мембраны для ионов калия и их диффузией по концентрационному градиенту.
В 1949 – 1952 гг. мембранно-ионную теорию модифицировали и экспериментально обосновали А. Ходжкин, А. Хаксли и др. Исследователям удалось найти замечательный объект — гигантский аксон кальмара, диаметром 1 мм , иннервирующий мышцы мантии. В такой аксон можно было легко вводить микроэлектрод, заменять внутреннее содержимое волокна искусственными растворами.
На основании проведенных опытов была сформулирована современная мембранная теория, основные положения которой следующие:
1. Мембрана клетки любого возбудимого образования в покое поляризована. При этом ее внутренняя поверхность заряжена отрицательно, а наружная — положительно;
2. Наличие электрических потенциалов в животных клетках обусловлено неравенством концентраций ионов Na + , K + , CI — , Ca 2+ внутри и вне клетки, а также их различной проницаемостью через мембрану.
3. В состоянии покоя внутри нервных и мышечных клеток концентрация K + в 30-40 раз выше, чем в наружном растворе. Концентрация Na + вне клетки в 10 –12 раз больше, чем внутри. Вне клетки больше также и ионов CI – .
4. В покое мембрана нервных клеток наиболее проницаема для ионов K + , менее для CI – и очень мало проницаема для ионов Na + (в 100 раз меньше, чем для K + ).
5. Для многих анионов органических кислот, присутствующих в цитоплазме, мембрана в покое не проницаема.
6. Благодаря преимущественной проницаемости мембраны для ионов К + в состоянии покоя, происходит их перемещение по концентрационному градиенту из клетки наружу.
7. В силу возникающего мембранного потенциала ионы K + по электрическому градиенту частично возвращаются обратно в клетку. Когда число выходящих из клетки ионов K + становится равным числу входящих в клетку, то на мембране устанавливается так называемый равновесный калиевый потенциал, обозначаемый Ек.
Читайте также: Как сшить ткань невидимый шов
II . Современные представления о структурно — функциональной организации мембран.
Клеточная (или плазматическая) мембрана представляет собой тонкую, гибкую и эластичную структуру толщиной всего 7,5-10 нм. Она состоит в основном из белков и липидов. Примерное соотношение ее компонентов таково: белки — 55%, фосфолипиды — 25%, холестерол — 13%, другие липиды — 4%, углеводы -3%.
Структурную основу клеточной мембраны (матрикс) составляет бимолекулярный слой фосфолипи дов. Фосфолипиды являются барьером для заряженных частиц и молекул водорастворимых веществ. Молекулы фосфолипидов мембраны состоят из двух частей, одна из которых несет заряд и гидрофильна, а другая, напротив, не заряжена и гидрофобна. Молекулы липидов в клеточной мембране ориентированы так, что их гидрофильные головки расположены снаружи, а гидрофобные хвостики – друг к другу. В толще клеточной мембраны молекулы фосфолипидов взаимодействуют незаряженными гидрофобными участками (они «спрятаны» от внутриклеточной и внеклеточной воды).
В липидном слое клеточных мембран содержится много холестерина. Обмен липидов, в отличие от белков, происходит медленнее. Однако возбуждение, например, нейронов мозга приводит к уменьшению содержания в них липидов. В частности, после длительной умственной работы, при утомлении количество фосфолипидов в нейронах уменьшается (может быть, с этим связана хорошая память у лиц напряженного умственного труда).
Состав мембранных липидов определяется средой обитания и характером питания. Так, увеличение растительных жиров в пищевом рационе поддерживает жидкое состояние липидов клеточных мембран и улучшает их функции. Избыток холестерина в мембранах увеличивает их микровязкость, ухудшает транспортные функции клеточной мембраны. Однако недостаток жирных кислот и холестерина в пище нарушает липидный состав и функции клеточных мембран.
В липидном бислое располагаются мембранные белки, большинство которых являются гликопротеинами. Различают три типа мембранных белков: 1) интегральные, которые пронизывают мембрану насквозь; 2) полуинтегральные и 3) периферические, которые выступают только над одной ее поверхностью, не достигая другой.
Многие интегральные белки формируют каналы (или поры), через которые во внутри- и внеклеточную жидкость могут диффундиро вать вода и водорастворимые вещества, особенно ионы. Благодаря избирательности действия каналов одни вещества диффундируют лучше других.
Другие интегральные белки функционируют как белки-переносчики, осуществляя транспорт веществ, для которых липидный бислой непроницаем. Иногда белки-переносчики действуют в направлении, противоположном диффузии, такой транспорт называют активным. Некоторые интегральные белки являются ферментами.
Интегральные белки мембраны могут служить также рецепторами для водорастворимых веществ, включая пептидные гормоны, поскольку мембрана для них непроницаема. Взаимодействие белка-рецептора с определенным лигандом приводит к конформационным изменениям молекулы белка, что, в свою очередь, стимулирует ферментативную активность внутриклеточного сегмента белковой молекулы или передачу сигнала от рецептора внутрь клетки с помощью вторичного посредника. Таким образом, интегральные белки, встроенные в клеточную мембрану, вовлекают ее в процесс передачи информации о внешней среде внутрь клетки.
Молекулы периферических мембранных белков часто бывают связаны с интегральными белками. Большинство периферических белков являются ферментами или играют роль диспетчера транспорта веществ через мембранные поры
Углеводы мембран. Клеточный гликокаликс. Углеводы мембран представлены главным образом в соединении с белками (гликопротеины) или жирами (гликолипиды). Большая часть интегральных белков на самом деле являются гликопротеинами, а примерно 1/10 часть всех липидных молекул мембраны — гликолипидами. Углеводные части мембранных структур почти всегда направлены наружу и выступают над поверхностью клетки. Обширную группу углеводных соединений составляют протеогликаны, состоящие из белка с отходящими от него боковыми углеводными цепями. Эти соединения обычно слабо связываются с мембраной и находятся на ее наружной поверхности. Таким образом, вся наружная поверхность клетки окружена углеводной оболочкой, которую называют гликокаликсом.
Углеводная оболочка клетки выполняет ряд важных функций: (1) большинство клеток способны отталкивать другие отрицательно заряженные частицы благодаря тому, что многие углеводные молекулы имеют отрицательный заряд; (2) гликокаликс соседних клеток скрепляет их друг с другом; (3) многие углеводные цепочки выполняют роль рецепторных молекул для связывания гормонов, включая инсулин.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом