
Некоторые из транспортных белков находятся в плазматической мембране, в то время как другие входят в состав мембраны клеточных органелл. Для поддержания определенного состава внутриклеточной среды и клеточных компартментов необходимо, чтобы транспортные белки обладали избирательностью по отношению к определенным метаболитам. В зависимости от характера процесса транспорта мембранные транспортные белки подразделяются на две группы: каналы и переносчики.
Канальные белки содержат поры, через которые при открытом канале с высокой скоростью проходят метаболиты. Белок-переносчик связывает метаболит, находящийся по одну сторону мембраны, после чего конформация его меняется, и метаболит высвобождается с другой стороны мембраны.
Существует несколько типов белков, образующих каналы в мембранах. Порины, присутствующие в клетках некоторых прокариот, в митохондриях и в нексусах, где соединяется цитоплазма соседних клеток, обеспечивают прохождение метаболитов в соответствии с размером их молекул.
К более селективным каналам относятся комплексы ядерных пор и каналы, обеспечивающие перемещение белков через мембрану эндоплазматического ретикулума. В статьях на сайте будут рассмотрены ионные каналы и аквапорины, через которые осуществляется селективный трансмембранный перенос ионов и молекул воды соответственно. К настоящему времени описано более 100 различных типов канальных белков. Все они обладают следующими характеристиками:
• селективностью по отношению к переносимому метаболиту,
• высокой скоростью переноса метаболита и
• воротным механизмом, регулирующим транспорт метаболитов.
Участок канального белка, через который осуществляется трансмембранный транспорт, называется порой канала. Канал может находиться в нескольких различных конфигурациях. Некоторые каналы состоят из одного белка, трансмембранные сегменты которого образуют пору. Другие существуют в виде олигомеров, состоящих из идентичных или разных субъединиц, которые вместе образуют пору.
Наконец существуют каналы, состоящие из двух и более субъединиц, каждая из которых сама образует пору. Часто комплексы олигомеров каналов регулируются или позиционируются на определенных мембранах с участием дополнительных субъединиц.
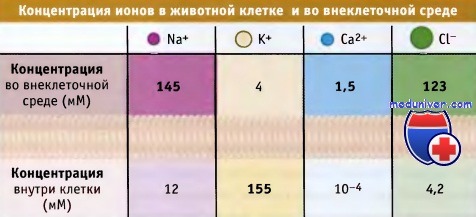
Большинство канальных белков обладают высокой селективностью в отношении определенных метаболитов, например ионов натрия (Na+), калия (К+), кальция (Са2+), хлорида (Сl-) или воды. Другие канальные белки являются неселективными катионными или анионными каналами. Как будет показано далее, при рассмотрении свойств специфических канальных белков, их поры характеризуются определенными структурными особенностями, т. е. являются селективным фильтром, который позволяет им проявлять избирательность по отношению к различным метаболитам.
![]()
Каналы и переносчики представляют собой два основных типа белков мембранного транспорта.
Растворенные метаболиты с максимальной скоростью диффундируют через пору каналов,
а переносчики связывают метаболит с одной стороны мембраны, затем претерпевают конформационные изменения и высвобождают его с другой стороны.
Этот процесс протекает значительно медленнее.
Направление потока ионов растворенных веществ через канал определяется электрохимическим градиентом. Иными словами, метаболиты движутся через канал в энергетически-выгодном направлении, т. е. по электрохимическому градиенту. Например, для покоящейся клетки, схематически изображенной на рисунке ниже, существует направленный в клетку ионный поток через Na+-, Са2+-, Cl—каналы, и противоположный, проходящий через К+-каналы.
Так как других источников энергии, кроме электрохимического градиента, в данном случае не используется, то такой тип транспорта называется пассивным. Канальные белки обеспечивают высокую скорость транспорта. Так, для ионных каналов скорость составляет 108 в 1 с, что близко к максимальной скорости диффузии ионов в водной среде.
Транспортные мембранные белки контролируют различные клеточные функции при участии воротного механизма. В основе этого механизма лежит индукция конформационных изменений в белке в ответ на воздействие определенного стимула. Эти изменения приводят к открытию или закрытию канала. Например, открытие и закрытие ионных каналов могут регулироваться лигандами, величиной электрического потенциала, механическими или температурными воздействиями. Активация каналов может происходить очень быстро, что делает их идеальным инструментом обработки поступающих в клетку сигналов.
Например, передача нервного импульса зависит от электрического заряда, который создается ионным каналом в мембране. Каналы также играют важную роль в регуляции объема клетки и внутриклеточного pH, в транспорте солей и воды через эпителиальные клетки, в закислении внутриклеточных органелл и во внеклеточной передаче сигналов.
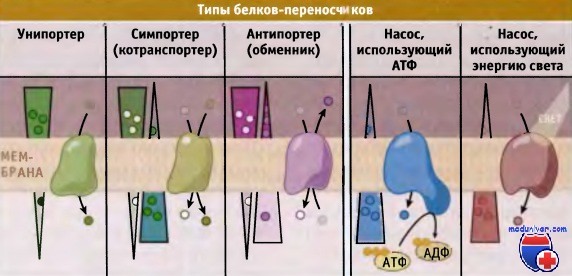
Транспортеры и насосы представляют собой два основных типа белков-переносчиков.
Транспортеры делятся на три типа, каждый из которых осуществляет один из типов переноса: унипорт, симпорт, или антипорт.
Показано направление электрохимического градиента через мембрану.
В зависимости от типа белка-переносчика транспорт может происходить или в направлении градиента (от высокой концентрации к низкой), или против него.
Читайте также: Какие функции выполняют ткани внутренней среды
При транспорте субстратов против градиента концентрации, белки-переносчики используют свободную энергию электрохимического градиента, АТФ или других источников. Поскольку при этом происходит потребление энергии, процесс носит название активный транспорт. Белки-переносчики подразделяются на две группы, транспортеры и насосы.
Для транспортировки субстратов через мембрану транспортеры используют энергию электрохимического градиента. Они подразделяются на унипортеры, симпортеры (или котранс-портеры) и антипортеры (обменники). Для энергозависимого транспорта метаболитов мембранные насосы используют энергию, например высвобождающуюся при гидролизе АТФ. По сравнению с канальными белками, переносчики характеризуются более медленной скоростью транспорта, порядка 1000 молекул в 1 с.
Различают два типа активного транспорта, первичный и вторичный. Белки-переносчики, осуществляющие первичный активный транспорт, в качестве источника энергии используют АТФ и транспортируют метаболиты против электрохимического градиента. Таким образом, они помогают поддерживать градиент концентрации метаболитов по обеим сторонам мембран клетки. Са2+-АТФаза и Na+/К+-АТФаза являются характерными примерами транспортных насосов, которые управляют процессом первичного активного транспорта.
Белки-переносчики, которые участвуют во вторичном активном транспорте, непосредственно не используют энергию АТФ. Вместо этого они используют свободную энергию электрохимического градиента, который возникает за счет первичного активного транспорта. Вторичный активный транспорт происходит с участием симпортеров и антипортеров. Механизмы транспорта метаболитов представлены на рисунке ниже.
В клетке все типы мембранных транспортных белков-каналы, транспортеры и насосы-функционируют совместно. Таким образом, работа какого-либо одного типа транспортного белка зависит от работы других. В данной главе мы рассмотрим несколько примеров такого взаимодействия транспортных белков. Например, ионный градиент по обеим сторонам мембраны поддерживается за счет сложных взаимодействий между различными типами транспортных белков. Для правильного функционирования эпителиальных клеток, например почек, слизистой кишечника и легкого, необходимо протекание транспортных процессов с участием различных ионов и метаболитов. Мы также рассмотрим роль нарушений процессов транспорта в развитии различных заболеваний.
![]()
Транспорт растворенных метаболитов через мембрану может быть пассивным или активным.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Питание клеток тканей транспорт
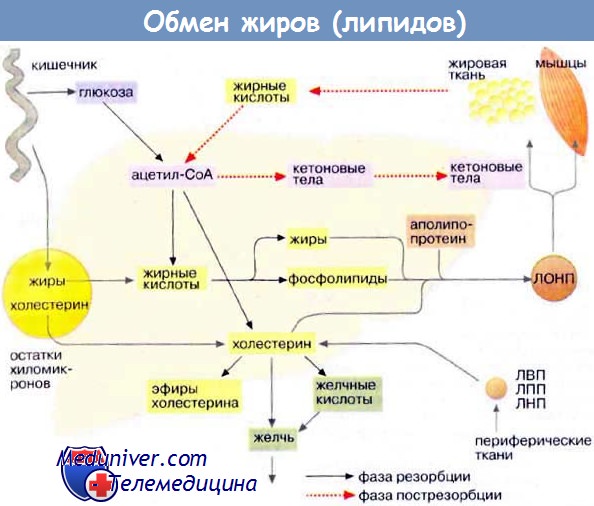
После всасывания в эпителий кишки свободные жирные кислоты и 2-моноглицериды вновь образуют триглицериды и вместе с фосфолипидами и холестеролом включаются в хиломикроны. Хиломикроны транспортируются с током лимфы через грудной проток в верхнюю полую вену, попадая таким образом в общий кровоток.
Внутри хиломикрона триглицериды гидролизуются липопротеинлипазой, что приводит к высвобождению жирных кислот на поверхности кровеносных капилляров в тканях. Это обусловливает транспорт жирных кислот в ткани и последующее образование остатков хиломикронов, обедненных триглицеридами. Эти остатки затем забирают эфиры холестерола из липопротеинов высокой плотности, и частицы быстро захватываются печенью. Данную систему транспорта жирных кислот пищевого происхождения называют экзогенной транспортной системой.
Также существует эндогенная транспортная система, предназначенная для внутриорганного транспорта жирных кислот, образованных в самом организме. Липиды транспортируются из печени в периферические ткани и обратно, а также переносятся из жировых депо к различным органам. Транспорт липидов от печени к периферическим тканям включает согласованные действия ЛПОНП, липопротеинов промежуточной плотности (ЛППП), липопротеинов низкой плотности (ЛПНП) и липопротеинов высокой плотности (ЛПВП). Частицы ЛПОНП подобно хиломикронам состоят из большого гидрофобного ядра, образованного триглицеридами и эфирами холестерола, и поверхностного липидного слоя, состоящего в основном из фосфолипидов и холестерола.
ЛПОНП синтезируются в печени, и отложение жира в периферических тканях является их основной функцией. После попадания в кровоток ЛПОНП подвергаются воздействию липопротеинлипазы, которая гидролизует триглицериды до свободных жирных кислот. Свободные жирные кислоты, происходящие из хиломикронов или ЛПОНП, могут быть использованы в качестве источников энергии, структурных компонентов фосфолипидных мембран или превращаться обратно в триглицериды и в таком виде сохраняться. Триглицериды хиломикронов и ЛПОНП также подвергаются гидролизу липазы печени.
Частицы ЛПОНП посредством гидролиза триглицеридов превращаются в более плотные, меньшие по размеру холестерол- и триглицерид-обогащенные остатки (ЛППП), которые удаляются из плазмы с помощью рецепторов печеночных липопротеинов или могут быть превращены в ЛПНП. ЛПНП являются основными липопротеинами-переносчиками холестерола.
Возврат липидов из периферических тканей в печень часто называют обратным транспортом холестерола. Частицы ЛПВП участвуют в этом процессе, забирая холестерол из тканей и других липопротеинов и перенося его в печень для последующей экскреции. Еще один вид транспортировки, существующей между органами, — перенос жирных кислот из жировых депо к органам для окисления.
Жирные кислоты, получаемые в основном в результате гидролиза триглицеридов жировой ткани, секретируются в плазму, где соединяются с альбумином. Связанные с альбумином жирные кислоты переносятся по градиенту концентрации в ткани с активным метаболизмом, где и используются преимущественно в качестве источников энергии.
Читайте также: Синяя ткань my time at portia
В течение последних 20 лет только несколько исследований были посвящены вопросу транспорта липидов в перинатальном периоде (результаты этих исследований в данном издании не представлены). Очевидна необходимость более детального изучения этой проблемы.
Жирные кислоты используются в качестве строительного материала в составе липидов клеточной стенки, в качестве источников энергии, а также откладываются «про запас» в виде триглицеридов преимущественно в жировой ткани. Некоторые омега-6 и омега-3 ДЦПНЖК являются предшественниками биологически активных метаболитов, используемых в сигнальной системе клеток, регуляции генов и других метаболически активных системах.
Вопрос о роли ДЦПНЖК ARA и DHA в процессе роста и развития ребенка является одним из важнейших вопросов в исследованиях, проводимых в области детской нутрициологии в течение последних двух десятилетий.
Липиды являются одними из основных компонентов клеточных мембран. Значительное количество исследований в области физиологии липидов посвящено двум жирным кислотам — ARA и DHA. ARA обнаружена в составе клеточных мембран всех структур организма человека; она является предшественником эйкозаноидов 2-й серии, лейкотриенов 3-й серии и других метаболитов, которые включены в сигнальные системы клеток и процесс генной регуляции. Исследования, посвященные DHA, часто указывают на ее структурную и функциональную роль в составе клеточных мембран.
Эта жирная кислота обнаружена в высокой концентрации в сером веществе головного мозга, а также в палочках и колбочках сетчатки. Исследования постепенного исключения из диеты животных омега-3 жирных кислот показали, что содержащие 22 атома углерода омега-6 ДЦПНЖК (например, 22:5 п—6) способны структурно, но не функционально заменить 22:6 n-3. При неадекватном уровне 22:6 n-3 в тканях выявляются нарушения зрения и познавательных способностей. Было показано, что изменение содержания 22:6 n-3 в тканях влияет на нейротрансмиттерную функцию, активность ионных каналов, сигнальные пути и экспрессию генов.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в оглавление раздела «физиология человека»
Питание клеток тканей транспорт
• Клетки эукариот обладают развитой системой внутренних структур, окруженных мембранами, которые называются органеллами
• Каждая органелла обладает уникальным составом (глико)протеинов и (глико)липидов и выполняет определенный набор функций
• Каждая органелла содержит один или несколько компартментов, ограниченных мембранами
• Органеллы выполняют свои функции автономно или в группах
• При эндоцитозе и экзоцитозе переносимые белки (белки карго) транспортируются между компартментами через транспортные везикулы, которые образуются отпочковыванием от поверхности органеллы и затем сливаются с мембраной-мишенью акцепторного компартмента
• Транспортные везикулы могут избирательно включать переносимый материал и исключать те компоненты, которые должны оставаться в органелле, из которой везикулы образовались
• Селективное включение в везикулы обеспечивается сигналами, присутствующими в первичной структуре белка или в углеводной структуре
• Транспортные везикулы содержат белки, которые направляют их к местам назначения и связывания. В дальнейшем везикулы сливаются с акцепторным участком мембраны
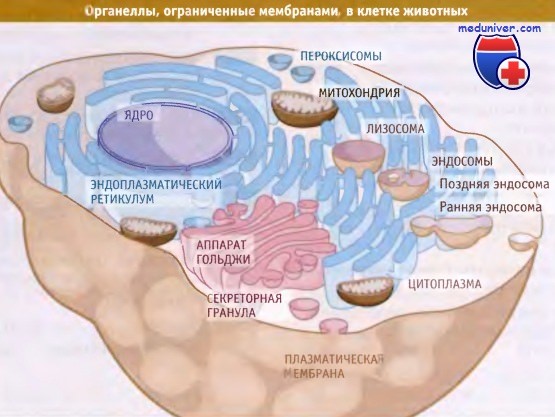
Компартменты, ограниченные мембранами в типичной клетке животных.
Одной из характерных особенностей эукариотической клетки является присутствие в ней развитой системы внутренних структур, окруженных мембранами, которые называются органеллы. Для клеток эукариот характерно присутствие мембран, разделяющих их внутреннее содержимое на функционально-различные компартменты, тогда как все клетки живых организмов обладают наружной двухслойной мембраной.
Одно из преимуществ компартментализации состоит в том, что клетка имеет возможность создавать необходимое окружение для выполнения функций, требующих определенного химического состава среды.
На проиллюстрировано строение и разнообразие органелл, имеющих мембрану, которые обычно присутствуют в эукариотической клетке (в данном случае в типичной животной клетке). Каждая органелла содержит один или несколько компартментов. Например, эндоплазматический ретикулум (ЭПР) представляет собой один компартмент; напротив, аппарат Гольджи состоит из нескольких окруженных мембранами компартментов, обладающих определенными биохимическими функциями.
Для митохондрии характерны два компартмента, матрикс и межмембранное пространство, содержащие набор определенных макромолекул.
Цитозоль может считаться одним компартментом, ограниченным плазматической мембраной и находящимся в контакте с наружной частью мембраны всех внутриклеточных органелл. Цитоплазма состоит из цитозоля и органелл. Аналогичным образом, нуклеоплазма ограничена внутренней ядерной мембраной.
Каждая органелла содержит уникальный набор белков (как мембранных, так и растворимых), липидов и других молекул, необходимых для выполнения ее функций. Некоторые липиды и белки ковалентно связаны с олигосахаридами. По мере роста и деления клеток должны синтезироваться их новые компоненты, необходимые для роста, деления и окончательного распределения внутриклеточного материала между двумя дочерними клетками. При дифференцировке и развитии клетки, а также в ответ на воздействие таких внешних факторов, как стресс, происходит синтез компонентов органелл.
Читайте также: Модели зимнего пальто из ткани
Однако компоненты не всегда образуются в той органелле, где они функционируют. Обычно различные макромолекулы образуются в сайтах, специально предназначенных для их синтеза. Например, большинство белков образуются на рибосомах цитозоля, который является оптимальной средой для функционирования рибосом и синтеза белка.
Возникает следующий вопрос: каким образом компоненты органелл попадают в места их функционирования? С начала 1970-х гг. этот вопрос являлся центральным в клеточной биологии. Как следует из рисунке ниже, существует, по крайней мере, восемь основных типов органелл, каждая из которых состоит из сотен или тысяч различных белков и липидов.
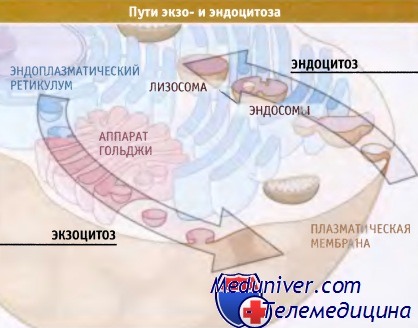
Экзоцитоз и эндоцитоз.
В экзоцитозе участвуют эндоплазматический ретикулум (включая ядерную оболочку)
и аппарат Гольджи (представлена одна стопка цистерн).
Эндоцитоз происходит с участием ранних и поздних эндосом и лизосом.
Все эти молекулы должны транспортироваться в органеллы, в которых они выполняют свои функции. Большинство белков образуется в цитозоле, и поэтому возникает вопрос: как они доставляются в соответствующие органеллы или же выходят из клетки, если принадлежат к секретируемым белкам? Во многих случаях ответом на этот вопрос служит наличие в молекуле белка специальных сигналов, обычно называемых сигналами сортировки или сигналами адресования. Они представляют собой короткие последовательности аминокислот, присутствующие в первичной структуре тех белков, которые должны быть локализованы не в цитозоле. Каждый адрес назначения белковой молекулы связан с одним или несколькими различными видами сигналов.
Сигналы сортировки узнаются специальными системами клетки по мере продвижения белка к месту назначения. Как показано на рисунке ниже, существует два основных транспортных механизма: экзоцитоз (или секреторный путь) и эндоцитоз, при котором материал (карго) транспортируется из клетки и в клетку соответственно.
Для всех вновь синтезированных белков, предназначенных для секреции из клетки, или для поступления в органеллы путем экзо- или эндоцитоза на мембране ЭПР существует общая точка входа. Сигналами к транслокации белка через мембрану ЭПР служат сигнальные последовательности. В настоящей главе мы рассмотрим сигналы сортировки, которые направляют белки к местам их назначения.
Находясь в ЭПР, белок не может транспортироваться через цитоплазму, и единственным путем его попадания в другие органеллы, окруженные мембранами, является везикулярный транспорт. Транспортные везикулы в основном состоят из белков и липидов и, как говорят, «отпочковываются» от мембраны. После того как везикула отпочковалась, она сливается со следующим компартментом, находящимся на ее пути. Компартмент, из которого возникла везикула, обычно называется донорский компартмент (или исходный компартмент), а компартмент назначения (или целевой) обычно называется акцепторный компартмент.
Транспортные везикулы прямым или непрямым путем переносят белки из ЭПР ко всем остальным компартментам на пути экзо- или эндоцитоза. При эндоцитозе везикулы образуются на плазматической мембране. Эти везикулы транспортируют заключенный в них материал в эндосомы, из которых образуются другие везикулы, переносящие материал в другие компартменты. Таким образом, состав транспортных везикул различается в зависимости от их происхождения и компартмента назначения.
Везикулярный транспорт создает проблему для органелл, с которыми обмениваются везикулы. Для нормального функционирования должен поддерживаться определенный внутренний состав органелл. Однако каким образом этого можно достичь, если везикулы все время изменяют этот состав? Масштаб проблемы становится очевидным при расчете эффективности транспорта. По пути эндоцитоза количество мембранных белков и липидов, эквивалентное их общему содержанию в плазматической мембране, способно транспортироваться через органеллы менее чем за час. Если сравнить со временем, необходимым для синтеза новой органеллы (обычно один день), то такая скорость впечатляет.
Решение этой проблемы связано с селективностью транспортного процесса. При отпочковывании в везикулу проходят только те белки, которые необходимо транспортировать. Резидентные белки органеллы в везикулу не попадают. Везикула удерживает эти белки и передает их следующей, находящейся на пути везикуле. Для поддержания гомеостаза между органеллами, по своей природе везикулярный транспорт всегда должен быть двунаправленным, т. е. компоненты донорского компартмента не должны непрерывно переноситься в акцепторный компартмент.
Механизм рециклирования возвращает некоторые компоненты везикул в донорский компартмент для их повторного использования. Поскольку селективность отбора белков несовершенна, специальный механизм утилизации возвращает назад резидентные белки, случайно попавшие в везикулу из донорского компартмента.
![]() При везикулярном транспорте везикула отпочковывается
При везикулярном транспорте везикула отпочковывается
от мембраны одного компартмента и сливается с мембраной другого компартмента. ![]() Пути транспорта белков в типичной животной клетке.
Пути транспорта белков в типичной животной клетке.
Почти все транспортные потоки могут осуществляться в двух направлениях.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом