
Развитие костной ткани на месте хряща протекает несколько сложнее, чем остеогистогенез, совершающийся непосредственно в мезенхиме. В этом случае развитию костной ткани предшествует образование хрящевой модели трубчатой кости, выполняющей опорную функцию на докостной стадии формирования скелета. Исходными клетками являются камбиальные клетки надхрящницы — адвентициальные. При подрастании к надхрящнице кровеносных сосудов и улучшении условий трофики и оксигенации эти клетки дифференцируются не в хондробласты, а в остеобласты, вырабатывающие межклеточное вещество ретикулофиброзной костной ткани. Они образуют подобие костной манжетки, окружающей хрящевую модель будущей трубчатой кости. Так возникает перихондральная костная ткань и надкостница. Окруженные костной тканью хрящевые клетки, утратившие связь с источником питания, подвергаются дегенерации. В возникшие полости дегенерирующего хряща из надкостницы врастают кровеносные сосуды с расположенными вокруг них камбиальными клетками. Некоторые из них превращаются в остеобласты, обусловливающие энхондральное развитие ретикулофиброзной костной ткани. Клетки, которые замуровываются в межклеточное вещество, дифференцируются в остеоциты, а периферически расположенные клетки — остеобласты — размножаются и продолжают синтез и секрецию компонентов межклеточного вещества. Все эти процессы первоначально протекают в середине хрящевой модели трубчатой кости (диафизе) и распространяются в проксимальном и дистальном направлениях.
В зоне контакта хрящевой и костной тканей можно выделить зоны неизмененного хряща, размножающихся хондроцитов, формирующих клеточные колонки, зону дегенерации и замещения хряща костной тканью. Зона размножающихся хрящевых клеток определяет зону роста будущей кости и важна для формирования вектора роста кости.
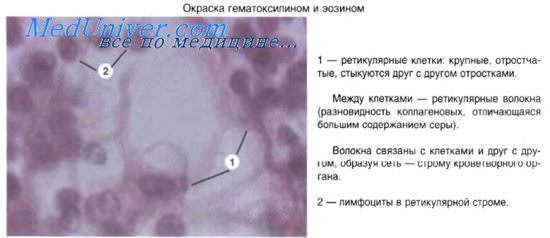
Одновременно с формированием ретикулофиброзной костной ткани, содержащей остеобласты и остеоциты, возникает другой гистогенетический тип клеток — остеокласты. Это крупные многоядерные (до 20-100 ядер) клетки размером до 100 мкм в диаметре являются производными стволовой кроветворной клетки. Цитоплазма остеокластов оксифильна со слабо развитой эндоплазматической сетью. Хорошо развит комплекс Гольджи. В цитоплазме много лизосом, содержащих кислую фосфатазу, коллагеназу, карбоангидразу и другие ферменты. Особенно много лизосом в той части цитоплазмы остеокластов, которая обращена к разрушаемой ткани. На этой поверхности имеются многочисленные выросты цитоплазмы, образующие подобие «щеточной (гофрированной) каемки». Остеокласты специализированы на «внеклеточной работе» лизосом: гидролитические ферменты из них выходят и резорбируют межклеточное вещество. Методами микрокиносъемки показано, что остеокласты подвергают деминерализации и разрушению оссеиновые волокна и аморфное вещество, а затем макрофаги фагоцитируют остатки органического субстрата. Остеокласты разрушают хрящевую ткань и ретикулофиброзную костную ткань, формируя каналы для врастающих сосудов и проникновения остеобластов.
Последующие стадии гистогенеза складываются из процессов новообразования костной ткани, ее разрушения остеокластами и перестройки — ремоделирования. Важным фактором гистогенеза пластинчатой костной ткани, входящей в состав трубчатой кости, является вектор роста кости. Он определяет направление движения остеокластов, следовательно, формирования каналов и врастание в них кровеносных сосудов (по вектору роста). Кровеносный сосуд, в свою очередь, определяет упорядоченное (концентрическое) расположение остеобластов вокруг себя. При этом остеобласты синтезируют межклеточное вещество, оссеиновые волокна которого упорядоченно (параллельно) располагаются возле остеобласта и при минерализации формируют костную пластинку, толщиной 3-10 мкм. Соседняя костная пластинка содержит оссеиновые фибриллы, которые располагаются под углом по отношению к первым.
На протяжении гистогенеза и всей возрастной динамики костной ткани в ней происходит непрерывная перестройка благодаря согласованной деятельности остеобластов и остеоцитов, образующих межклеточное вещество, а также остеокластов, разрушающих костную ткань, что необходимо для процессов ее самообновления. Так происходит смена генераций костных пластинок и формирующихся структурно-функциональных единиц — остеонов, достигается упорядоченность расположения последних, следовательно, высокая механическая прочность костной ткани и кости как органа (см. кость).
Дентиноидная костная ткань отличается отсутствием тел костных клеток в толще межклеточного вещества. Дентин — это вещество, состоящее из коллагеновых волокон и основного аморфного вещества, пропитанного минеральными солями. Образующие дентин зуба клетки — одонтобласты (точнее — их ядросодержащая часть) — расположены вне дентина в пульпе зуба. Дентин пронизан дентинными канальцами, в которых проходят отростки одонтобластов. Сходное строение имеет цемент зуба.
Ретикулофиброзная (грубоволокнистая) костная ткань характеризуется беспорядочным расположением оссеиновых фибрилл в виде толстых, плотных пучков волокон и основного аморфного вещества. Такая костная ткань образует кости в зародышевом и раннем постнатальном периодах. У взрослого человека она сохраняется лишь на месте прикрепления сухожилии к кости, в зарастающих швах черепа, а также в составе тканевого регенерата на месте переломов костей.
Пластинчатая костная ткань отличается упорядоченным расположением оссеиновых фибрилл в составе костных пластинок. Последние образуют расположенные один за другим слои пропитанного солями кальция фибрилл, образованных остеобластами. Слои имеют толщину от 3-7 до нескольких сотен микрометров. Каждая костная пластинка состоит из параллельно ориентированных тонких оссеиновых (коллагеновых) волокон (коллаген 1-го типа). Но коллагеновые волокна двух прилежащих друг к другу костных пластинок ориентированы под разными углами. Костная пластинка соединяется с соседней пластинкой коллагеновыми фибриллами. Так создается прочная волокнистая основа кости. Костные пластинки располагаются концентрически вокруг сосудов, то есть формируют остеоны — структурно-функциональные единицы пластинчатой кости как органа. Кроме этого существуют наружные и внутренние окружающие и вставочные пластинки трубчатой кости (см. ниже).
Читайте также: Темно синяя пальтовая ткань
Регенерация. В регенерации костной ткани участвуют детерминированные остеогенные элементы в составе надкостницы, механоциты костного мозга, которые размножаются и дифференцируются в остеобласты. Продуцируя межклеточное вещество, остеобласты дифференцируются в остеоциты и образуют ретикулофиброзную костную ткань. Кроме того, адвентициальные клетки волокнистой соединительной ткани надкостницы также принимают участие в регенерации костной ткани. Однако дифференцировка их во многом зависит от микроокружения, внетканевых и внеорганных факторов (например, от репозиции отломков, неподвижности отломков, оксигенации места перелома и др.).
Дифференцировка адвентициальных клеток возможна в трех направлениях: остеогенном, хондрогенном, фибробластическом. Этим определяется соотношение различных видов тканей в регенерате. При преимущественно остеобластическом гистогенезе формируется ретикулофиброзная костная ткань, которая постепенно ремоделируется с образованием костной ткани, напоминающей по своему строению пластинчатую.
Текст книги «Общая и частная гистология»

Автор книги: Рудольф Самусев
Жанр: Медицина, Наука и Образование
Текущая страница: 8 (всего у книги 25 страниц) [доступный отрывок для чтения: 6 страниц]
Прямой (внутримембранный) остеогистогенез (рис. 6.5, А):

А – из мезенхимы: 1 – мезенхима; 2 – остеобласты; 3 – остеоциты; 4 – остеокласт; 5 – остеоид; 6 – кровеносные сосуды (×130);
– характерен для развития грубоволокнистой костной ткани;
– этим способом образуются плоские кости черепа, ключицы, дистальные фаланги пальцев;
– мезенхимные клетки группируются в остеогенные островки, которые являются первичными центрами окостенения;
– клетки островков дифференцируются в остеобласты, начинающие вырабатывать остеоид (органический матрикс кости);
– остеоид минерализуется и замуровывает остеобласты, которые превращаются в остеоциты;
– формируются трабекулы, образующие губчатую кость. Непрямой (энхондральный) остеогистогенез (рис. 6.5, Б):

Б – на месте хряща: 1 – зона пузырчатого хряща; 2 – зона столбчатого хряща; 3 – обызвествленный хрящ; 4 – костная манжетка; 5 – надкостница; 6 – костно-хрящевое соединение (×30).
– из мезенхимы образуются модели трубчатых костей, состоящие из гиалинового хряща;
– первичный центр окостенения (перихондральный) образуется в результате усиленного окостенения надхрящницы и образования в ней грубоволокнистой костной ткани (костная манжетка) в области диафиза;
– в результате этого нарушается питание хряща, возникают дистрофия и дегенерация хондроцитов с образованием в хряще полостей;
– сосуды прорастают из надкостницы в образовавшиеся полости, а вместе с ними остеогенные клетки и остеобласты;
– дифференцировка и продуктивная деятельность их приводит к образованию кости (энхондральное окостенение). На препаратах различимы компоненты комплекса: «кальцинированный хрящ (базофилен) – кальцинированная кость (оксифильна)»;
– костно-мозговая полость образуется с помощью остеокластов. Образованная ранее костная манжетка растет по направлению к эпифизам, где формируются вторичные центры окостенения;
– хрящевая ткань сохраняется только на суставных поверхностях и в метафизах в виде метаэпифизарной хрящевой пластинки;
– происходит перестройка грубоволокнистой кости в пластинчатую.
Рост трубчатых костей в длину происходит за счет эпифизарной хрящевой пластинки, состоящей из нескольких зон:
– резервная зона покоящегося хряща расположена в эпифизарной части пластинки, состоит из гиалинового хряща, содержащего небольшие хондроциты;
– зона размножения (пролиферативная) состоит из многочисленных делящихся хрящевых клеток, расположенных в виде «монетных» столбиков;
– зона гипертрофии и созревания хряща состоит из крупных вакуолизированных, дегенеративно измененных хондроцитов, прекративших митозы;
– зона кальцификации хряща, где происходят минерализация хрящевого матрикса, гибель хондроцитов и оссификация;
– зона окостенения, где на месте обызвествленного хряща формируется костная ткань.
Перестройка костной ткани:
– кость является динамичной структурой с постоянно изменяющимися формой и внутренней организацией;
– участки кости, испытывающие сжатие, подвергаются резорбции;
– в области приложения тянущих усилий образуется новая костная ткань;
– костные клетки чувствительны к пьезоэлектрическим эффектам, возникающим при деформации кости, что влияет на интенсивность остеогенеза.
– при переломе нарушается кровоснабжение и остеоциты гибнут;
– отмирающая кость подвергается активной резорбции остеокластами;
– между концами отломков формируется костная мозоль вследствие интенсивного размножения остеогенных клеток надкостницы;
– рост сосудов отстает от размножения клеток, поэтому сначала формируется хрящ, который замещается губчатой костью, перестраивающейся затем в компактную. Восстанавливается первоначальная конфигурация кости.
Регуляция роста и регенерации кости:
– паратареоидшый гормон (ПТГ) повышает содержание Са 2 + в крови за счет высвобождения его из костей, снижения экскреции его почками и стимуляции перехода витамина D в кальцитриол;
– кальцитонин – антагонист ПТГ, снижает содержание Са 2+ в крови за счет ингибирования резорбции костей остеокластами;
– активный метаболит витамина D кальцитриол усиливает всасывание Са 2+ в желудочно-кишечном тракте. При дефиците витамина D развивается рахит у детей и остеомаляция у взрослых;
– витамин C необходим для образования коллагена. При его дефиците замедляются рост костей и заживление переломов;
– витамин А поддерживает образование и рост кости. Недостаток его тормозит остеогенез и рост костей, избыток вызывает зарастание эпифизарных хрящевых пластинок и замедление роста кости в длину;
– глюкокортикоиды ухудшают всасывание Са 2+ в тонкой кишке;
– при недостатке эстрогенов развивается остеопороз.
Тесты и вопросы для самоконтроля
1. У ребенка, страдающего рахитом, наблюдаются искривление и размягчение костей конечностей. Какой этап костеобразования нарушен?
2. В организме недостаточно витамина С. Какой процесс в развитии костной ткани будет нарушен?
3. Какой патологический процесс будет наблюдаться в костной ткани при избытке паратиреоидного гормона?
4. При рентгеноскопии костей конечностей у больного не обнаружена эпифизарная пластинка роста. Какого возраста достиг больной?
5. С какой особенностью состава костной ткани связана легкость переломов у лиц пожилого возраста и редкость переломов у детей?
Выберите один или несколько правильных ответов:
6. Хондробласты. Верны все утверждения, кроме:
а) располагаются в надхрящнице;
б) участвуют в аппозиционном росте хряща;
г) участвуют в резорбции хряща.
7. Эластический хрящ. Верны все утверждения, кроме:
а) снаружи покрыт надхрящницей;
б) входит в состав слуховой трубы;
в) с возрастом обызвествляется;
г) содержит эластические и коллагеновые волокна.
8. Какой костной тканью образованы заросшие черепные швы?
9. Вставочные пластинки в диафизе трубчатых костей – это:
а) материал для образования общих пластин;
б) материал для образования остеонов;
в) оставшиеся части концентрических пластин старых остеонов;
г) часть вновь формирующихся остеонов.
а) локализуются преимущественно в местах перестройки кости;
б) хорошо развиты гранулярная эндоплазматическая сеть и комплекс Гольджи;
в) характерна невысокая активность щелочной фосфатазы;
11. Развитие кости на месте хряща:
а) это способ образования плоских костей;
б) костная манжета формируется вокруг эпифиза;
в) сопровождается гипертрофией хондроцитов в центральной части хряща;
Читайте также: Код окпд 2 фильтровальная ткань
г) остеогенные клетки проникают в полости центральной части хряща.
12. Пластинчатая костная ткань. Верно все, кроме:
а) образует компактное и губчатое вещество костей скелета;
б) формируется путем образования новых слоев на поверхности кости;
в) коллагеновые волокна построены из коллагена III типа;
г) коллагеновые волокна в пределах костной пластинки ориентированы упорядоченно;
д) каналы остеонов содержат кровеносные сосуды.
Ответы
2. Образование коллагеновых волокон.
5. С различием в степени минерализации костей.
Глава 7
Мышечные ткани
Мышечные ткани (textus musculares) представляют собой группу тканей различного происхождения и строения, объединенных на основании общего признака – выраженной сократительной способности, благодаря которой они могут выполнять свою основную функцию – перемещение тела и его частей в пространстве.
Эти ткани классифицируют по морфологии и функции:
• поперечнополосатая мышечная ткань имеет регулярную поперечнополосатую исчерченность и включает в себя скелетную мускулатуру и сердечную мышцу;
• гладкая мышечная ткань не имеет поперечнополосатой исчерченности и расположена в стенках внутренних органов и кровеносных сосудов;
• сокращение скелетной мускулатуры произвольное (подконтрольно сознанию), в то время как сокращение сердечной и гладкой мускулатуры непроизвольное.
7.1. Поперечнополосатая [скелетная] мышечная ткань
Поперечнополосатая мышечная ткань (textus muscularis striatus [skeletalis]) является самой распространенной мышечной тканью человека. У взрослых мужчин она составляет более 40 % от массы тела, у женщин – 35 %, у детей – около 25 %; при старении ее относительная масса снижается до 28–30 %. Помимо скелетных мышц, обеспечивающих поддержание позы и перемещение тела и его частей в пространстве, она образует глазодвигательные мышцы, мышцы стенки полости рта, языка, глотки, гортани, верхней трети пищевода, наружный сфинктер прямой кишки.

Рис. 7.1. Скелетная мышечная ткань языка. ×400.
1 – эндомизий; 2 – перемизий; 3 – мышечные волокна (срезаны поперечно); 4 – ядра мышечных волокон; 5 – мышечные волокна с поперечной исчерченностью (срезаны продольно).
– состоит из мышечных волокон, окруженных соединительной тканью;
– соединительно-тканные оболочки связаны с мышцами на трех главных уровнях организации:
– эпимизий окружает всю мышцу;
– перимизий охватывает каждый мышечный пучок, состоящий из 20—100 мышечных волокон;
– эндомизий окружает каждое волокно.
– поперечнополосатая мышечная ткань гетерогенна в том смысле, что состоит из трех разных типов волокон;
– типы волокон различаются по содержанию пигмента миоглобина, гликогена, количеству митохондрий и концентрации различных ферментов, способу энергообеспечения. Различают: тип I (красные), тип 11В (белые) и тип IIA (промежуточные).
Красные волокна содержат большое количество пигмента миоглобина, много крупных митохондрий и липидных включений, богаты окислительными ферментами; это тонические, устойчивые к утомлению, с небольшой силой сокращения окислительные волокна;
– характеризуются малым диаметром, относительно тонкими миофибриллами, богатым кровоснабжением;
– получили название «медленные волокна», поскольку легко стимулируются, однако скорость их сокращения низкая, они приспособлены к медленным повторным сокращениям;
– демонстрируют слабое цитохимическое окрашивание на АТФазу;
– энергообеспечение аэробного характера;
– преобладают в мышцах, выполняющих длительные тонические нагрузки (мышцы туловища).
Белые волокна содержат меньше миоглобина и митохондрий, чем красные;
– характеризуются большим диаметром, крупными и сильными миофибриллами, значительным количеством гликогена, сравнительно слабым кровоснабжением;
– бедны ферментами окисления, но богаты фосфорилазами;
– имеют высокую скорость сокращения и приспособлены к быстрым кратковременным усилиям; это тетанические, с большой силой сокращения, легко утомляющиеся гликолитические волокна;
– демонстрируют заметное цитохимическое окрашивание на АТФазу;
– имеют анаэробное энергообеспечение;
– преобладают в мышцах, выполняющих быстрые движения (мышцы конечностей).
Промежуточные волокна обладают характеристиками, промежуточными между уже описанными для красных и белых волокон;
– быстрые, устойчивые к утомлению, с большой силой сокращения окислительно-гликолитические волокна.
Иннервация определяет дифференцировку типов волокон: красные волокна имеют небольшое количество нервных двигательных окончаний, белые – значительно больше.
Скелетные мышцы человека являются смешанными, т. е. содержат волокна различных типов, которые распределены в них мозаично.
Соотношение красных и белых волокон в мышцах человека индивидуально, предопределено генетически и почти не меняется с возрастом.
Волокна скелетных мыщц – длинные, многоядерные симпласты, образованные миофибриллами. Каждая миофибрилла имеет поперечные полоски (А-диски и I-диски), которые повторяются и формируют характерный узор (см. рис. 7.1);
– на гистологических срезах А-диск анизотропен и обычными гистологическими красителями окрашивается в темный цвет;
– I-диск изотропен и на обычных гистологических препаратах окрашен светло;
– Z-линия, или телофрагма, – это плотная область, которая делит I-диск пополам;
– H-полоска, или светлая зона, рассекает A-диск и имеет в центре темную M-линию, или мезофрагму;
– участок миофибриллы между двумя соседними линиями Z называется миомером, или саркомером, который является структурно-функциональной единицей поперечнополосатого мышечного волокна;
– в расслабленной мышце длина саркомера составляет около 2–3 мкм, при сокращении мышцы саркомер укорачивается до 1,5 мкм, т. е. почти вдвое;
– на электронной микрофотографии видно, что волокно скелетной мышцы окружено плазматической мембраной, называемой сарколеммой;
– имеет много ядер, лежащих на периферии сразу под сарколеммой;
– цитоплазму мышечного волокна называют саркоплазмой;
– содержит простирающиеся по всей длине волокна миофибриллы;
– миофибриллы имеют вид нитей диаметром 1–2 мкм и длиной, сопоставимой с протяженностью волокна. Их количество в отдельном волокне варьирует в широких пределах – от нескольких десятков до 2000 и более;
– миофибриллы обладают собственной поперечной исчерченностью, причем в мышечном волокне они располагаются так упорядоченно, что А– и I-диски одних миофибрилл точно совпадают с аналогичными дисками других, обусловливая поперечную исчерченность всего волокна.
Саркоплазматическая сеть – это модифицированная гладкая эндоплазматическая сеть;
– окружает миофиламенты и образует сеть вокруг каждой миофибриллы;
– на границе А– и I-дисков формирует расширенные терминальные цистерны, которые проходят вокруг миофибрилл;
– регулирует мышечное сокращение путем аккумулирования Ca 2+ (ведет к расслаблению) или освобождения Ca 2+ (ведет к сокращению миофибрилл).
Триады – характерная черта строения скелетных мышечных волокон:
– будучи расположенной на границе А– и I-дисков, каждая триада состоит из трубочки (Т-трубочка) и двух терминальных цистерн саркоплазматической сети;
– Т-трубочка – это глубокая инвагинация сарколеммы (плазматической мембраны) внутрь мышечного волокна;
– между Т-трубочкой и терминальными цистернами находятся регулярно расположенные уплотнения (соединительные ножки), представляющие собой зоны низкого электрического сопротивления;
– Т-трубочки быстро проводят импульсы с наружной поверхности волокна в более глубокие его области, которые запускают механизм выделения Ca 2 + из терминальных цистерн в саркоплазму, что способствует сокращению миофиламентов.
Читайте также: Мембранные ткани характеристики свойства
Миофибриллы состоят из продольно расположенных цилиндрических пучков белковых нитей – миофиламентов;
– лежат параллельно продольной оси волокна и простираются на всю его длину;
– имеют поперечную исчерченность;
– находятся в определенном положении друг к другу благодаря промежуточным филаментам, состоящим из белков десмина и виментина;
– каждая окружена саркоплазматической сетью. Миофиламенты расположены в миофибрилле строго упорядоченно и подразделяются на тонкие и толстые филаменты;
– I-диск содержит только тонкие филаменты, которые закрепляются на плотной Z-линии;
– A-диск состоит из толстых и тонких филаментов (это обусловливает ее двойное лучепреломление, или анизотропию);
– соотношение тонких и толстых филаментов 6: 1;
– М-линия образована поперечными соединениями из белка меромиозина между толстыми миофиламентами А-диска;
– Н-полоска состоит только из толстых филаментов. Толстые миофиламенты имеют диаметр 10–20 нм и длину 1,5–2,0 мкм;
– состоят из множества молекул фибриллярного белка миозина, организованных так, что они формируют палочковидную структуру (см. рис. 7.1);
– каждая молекула миозина имеет длину 150 нм и толщину 2 нм и выглядит похожей на клюшку для гольфа с двумя головками – глобулярными выступами, имеющими специфический участок для связывания АТФ;
– гидролиз АТФ происходит в головках, которые являются также местом связывания с актином;
– молекула миозина состоит из двух идентичных тяжелых цепочек и двух пар легких цепочек;
– тяжелые цепочки ферментативно расщепляются, формируя два фрагмента – легкий и тяжелый меромиозин;
– легкий меромиозин составляет большую часть палочковидного участка молекулы, в то время как тяжелый меромиозин представляет глобулярную головку и небольшую часть (шейку) палочки;
– молекула миозина может сгибаться, как на шарнирах, в месте соединения тяжелого меромиозина с легким и в области прикрепления головки;
– перемычки, видимые между тонкими и толстыми миофиламентами, сформированы головками молекул миозина плюс небольшая часть палочковидного участка.
Тонкие миофиламенты состоят из F-актина, который формируется при полимеризации мономеров G-актина, или глобулярного актина, и двух регуляторных белков – тропонина и тропомиозина;
– имеют в диаметре 5–7 нм и в длину 1 мкм;
– каждый актиновый филамент состоит из двух F-нитей в форме двойной спирали толщиной 6–7 нм и вариабельной длины;
– нитевидная молекула тропомиозина лежит в углублении актиновой двойной спирали;
– тропонин распределен вдоль тонких филаментов с интервалами 40 нм.
Тропонин – глобулярный белковый комплекс, состоящий из трех субъединиц:
– TnC, которая связывает Ca 2 +;
– TnT, которая прикрепляется к тропомиозину;
– TnI, которая ингибирует (предотвращает) взаимодействие актина и миозина.
В покое миозиновые головки, с которыми связаны молекулы АТФ, не способны взаимодействовать со специальными (актиновыми) центрами на молекуле актина, так как последние прикрыты комплексом тропонин-тропомиозин (КТТ).
Сокращение миофибриллы инициируется, когда Ca 2 + присоединяются к TnC-единице тропонина, воздействуя на КТТ;
– возникающее изменение конформации КТТ и смещение молекулы связанного с ним тропомиозина демаскирует (открывает) активные центры на молекуле актина, с которыми связываются миозиновые головки, образуя поперечные мостики;
– одновременно с этим головка молекулы миозина расщепляет АТФ на АДФ с выделением энергии;
– выделенная энергая способствует движению головки миозина и связанные с ней тонкие филаменты;
– таким образом, тонкий филамент скользит вдоль толстого филамента, когда происходит сокращение.
Гипотеза скользящих нитей (гипотеза Хаксли) предполагает, что сами филаменты не изменяют свою длину, а скользят друг вдоль друга, увеличивая степень перекрытия между ними;
– скольжение является результатом повторяющихся соединений и разъединений головок молекул миозина и соседних актиновых филаментов;
– результат: длина саркомера уменьшается практически вдвое;
– тонкие филаменты скользят, проникая глубже в А-полос-ку, но сама А-полоска остается неизменной длины;
– полоска I и полоска H уменьшаются в размере, когда линии Z приближаются к концам полосок A;
– расслабление после мышечного сокращения происходит в результате снижения концентрации Ca 2+ в области саркомера, которое вызывает отщепление Ca 2 +от TnC-субъединицы тропонина и возвращение тропонина в первоначальное состояние. Нити тропомиозина при этом вновь закрывают активные центры на молекулах актина, что обусловливает прекращение циклического образования мостиков между толстыми и тонкими филаментами.
Опорный аппарат мышечного волокна включает особые элементы цитоскелета (тело– и мезофрагма), а также связанную с ними сарколемму и базальную мембрану.
Телофрагма (линия Z) – область прикрепления тонких миофиламентов двух соседних саркомеров, ее ширина 30—100 нм;
– представляет из себя сложную трехмерную решетку из особых тонких нитей (Z-филаментов), идущих под углом 45° к оси саркомера и связывающих тонкие миофиламенты друг с другом. В состав линий Z входят также ряд белков: α-актинин, филамин, Z-белок.
Мезофрагма (линия M) – плотная линия шириной 75–85 нм, является областью закрепления толстых (миозиновых) филаментов в саркомере. Образована центральными участками миозиновых филаментов, связанных друг с другом мостиками, состоящими из белков миомезина, креатинкиназы и М-белка.
Титин (коннектин) – белок с эластическими свойствами, нити которого присоединены к толстым филаментам по всей их длине и, продолжаясь в I-диски, прикрепляют концы толстых филаментов к линиям Z. Нити титина препятствуют перерастяжению мышцы.
Небулин – белок в виде нитей, расположенных по всей ширине I-диска параллельно тонким филаментам, с которыми он связан, обеспечивая их механическую стабилизацию.
Промежуточные филаменты из белка десмина (толщиной 8– 10 нм) связывают, с одной стороны, соседние телофрагмы одной миофибриллы, с другой – мезофрагмы, а также телофрагмы соседних миофибрилл друг с другом;
– такие же филаменты прикрепляют телофрагмы к сарколемме и элементам системы Т-трубочек и саркоплазматической сети.
Костамеры – кольца из белка винкулина, охватывающие изнутри мышечное волокно и расположенные перпендикулярно к его длинной оси. Представляют собой участки непосредственного соединения между сарколеммой и подлежащими I-дисками миофибрилл. Помимо винкулина, в костамерах имеются другие белки, связанные с цитоскелетом: талин, спектрин, α-актинин.
Энергетический аппарат мышечных волокон представлен митохондриями, располагающимися в виде цепочек под сарколеммой и между миофибриллами, трофическими включениями (гликоген, липиды), которые находятся между миофибриллами по всей толщине миосимпласта, а также миоглобином.
Миоглобин – железосодержащий кислородсвязывающий пигмент мышечного волокна, придающий ему красный цвет. Его способность к связыванию кислорода способствует повышению активности процессов окислительного фосфорилирования, лежащего в основе энергетического обеспечивания мышечного сокращения.
Синтетический аппарат мышечного волокна представлен свободными рибосомами и полирибосомами, цистернами гранулярной эндоплазматической сети и комплексом Гольджи; аппарат внутриклеточного переваривания – лизосомами, необходимыми для постоянно протекающего процесса обновления структурных компонентов мышцы.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом