
Около 40% массы тела человека составляют скелегные мышцы, и, вероятно, 10% приходится на долю гладких мышц и мышцы сердца. Некоторые из основных принципов сокращения являются общими для разных типов мышц.
На рисунке показана схема организации скелетной мышцы. Все скелетные мышцы содержат множество волокон диаметром от 10 до 80 мкм. На рисунке также показано, что каждое из этих волокон состоит из последовательно соединенных более мелких элементов.
В большинстве скелетных мышц каждое волокно вытянуто во всю длину мышцы и, за исключением примерно 2%, обычно иннервируется лишь одним нервным окончанием вблизи середины волокна.
Сарколемма. Сарколеммой называют клеточную мембрану мышечного волокна. Она состоит из собственно мембраны клетки, называемой плазматической мембраной, и наружного покрытия из тонкого слоя полисахаридного материала, который содержит множество тонких коллагеновых нитей. На концах мышечного волокна этот поверхностный слой сарколеммы сливается с сухожильными волокнами. Сухожильные волокна, в свою очередь, собираются в пучки и формируют мышечные сухожилия, вплетающиеся в кости.
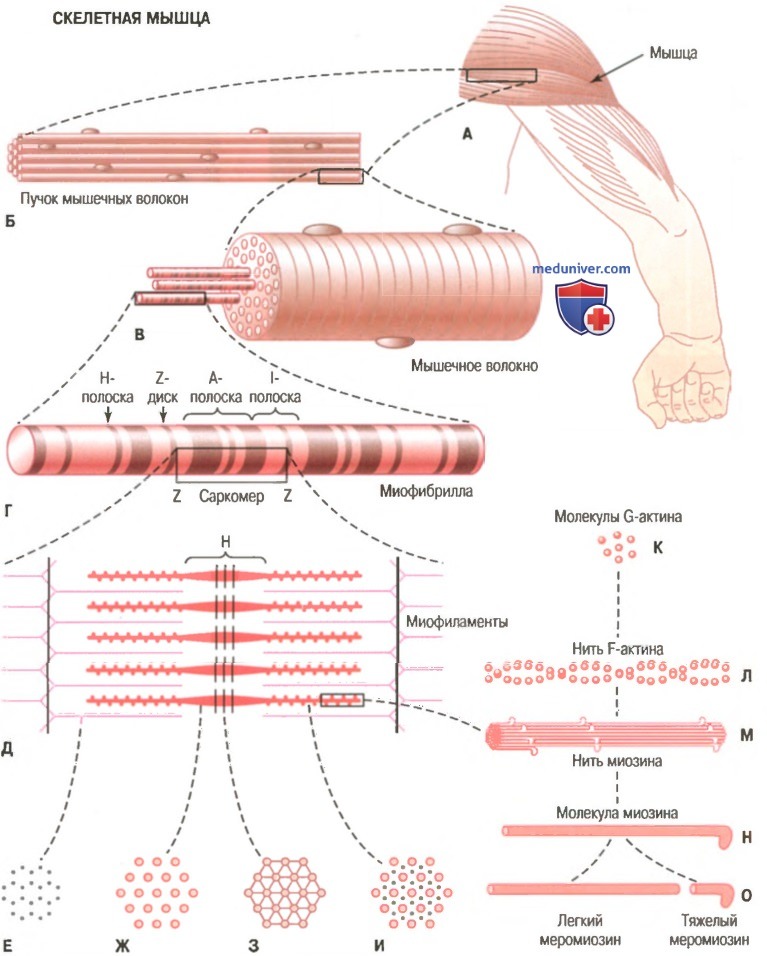
Организация скелетной мышцы от макроскопического до молекулярного уровня. Е, Ж 3, И — поперечное сечение на указанном уровне.
Миофибриллы. Нити актина и миозина. Каждое мышечное волокно содержит от нескольких сотен до нескольких тысяч миофибрилл, которые представлены на поперечном срезе в виде небольших светлых точек. Каждая миофибрилла собрана примерно из 1500 смежных нитей миозина и 3000 нитей актина, представляющих собой большие полимеризованные белковые молекулы, ответственные за мышечное сокращение. Они видны в продольном сечении на электронной микрофотографии. Толстые нити на схемах — миозиновые, тонкие — актиновые.
Обратите внимание, что на рисунке актиновые и миозиновые нити частично перекрываются, в связи с чем миофибриллы имеют чередующиеся светлые и темные полоски. Светлые полоски содержат только актиновые нити, их называют I-полосками, поскольку они изотропны в поляризованном свете. Темные полоски содержат миозиновые нити, а также концы актиновых нитей, которые частично накладываются на миозиновые нити. Эти полоски называют А-полосками, поскольку они анизотропны в поляризованном свете. На рисунке видны также маленькие выступы по сторонам миозиновых нитей. Это так называемые поперечные мостики. Их взаимодействие с актиновыми нитями лежит в основе сокращения мыщц.
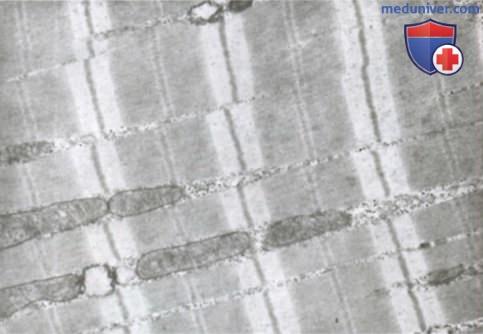
Электронная микрофотография миофибриллы, демонстрирующая детальную организацию актиновых и миозиновых нитей. Видны митохондрии, лежащие между миофибриллами.
На рисунке показано также, что концы актиновых нитей прикреплены к так называемому Z-диску (Z-мембране). Отсюда актиновые нити распространяются в обе стороны, проходя между миозиновыми нитями. Z-диск, состоящий из нитевидных белков, отличающихся от актиновых и миозиновых нитей, проходит в поперечном направлении через миофибриллу, а также через все мышечное волокно, переходя от одной миофибриллы к другой, скрепляя их между собой. В связи с этим целое мышечное волокно, как и отдельные миофибриллы, имеет светлые и темные полоски. Эти полоски придают скелетным и сердечной мышцам характерный полосатый вид.
Часть миофибриллы (или целого мышечного волокна), расположенную между двумя последовательными Z-дисками, называют саркомером. В сокращенном мышечном волокне длина саркомера равна примерно 2 мкм. При этой длине актиновые нити полностью перекрывают миозиновые, и кончики актиновых нитей начинают перекрывать друг друга. Мы увидим далее, что при этой длине мышца способна генерировать максимальную силу сокращения.
Что удерживает актиновые и миозиновые нити на месте? Удерживать актиновые и миозиновые нити рядом довольно трудно. Это обеспечивается с помощью большого числа нитевидных молекул белка, называемого тайтином. Каждая молекула тайтина имеет молекулярную массу около 3000000, что делает ее одной из самых крупных белковых молекул в организме. Кроме того, поскольку молекула тайтина волокнистая, она очень упруга. Упругие молекулы тайтина образуют каркас, удерживающий актиновые и миозиновые нити в положении, обеспечивающем нормальную работу сократительного аппарата саркомера. Это позволяет полагать, что сама молекула тайтина действует как матрица для начального формирования участков сократительных нитей саркомера, особенно миозиновых нитей.
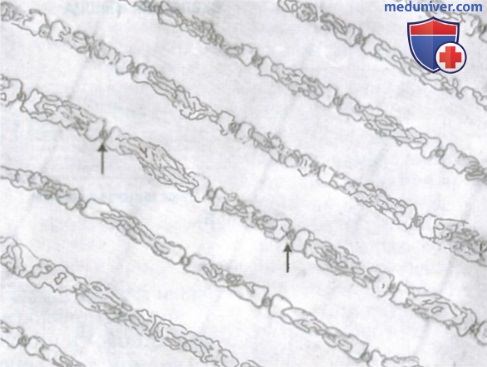
Саркоплазматический ретикулум во внутриклеточных пространствах между миофибриллами с системой продольных трубочек, параллельных миофибриллам. Показаны также в поперечном сечении Т-трубочки (стрелки), которые связаны с внеклеточным пространством и отвечают за проведение электрического сигнала в центр мышечного волокна.
Саркоплазма. В каждом мышечном волокне многочисленные миофибриллы расположены рядом, бок о бок. Пространство между ними заполнено внутриклеточной жидкостью, называемой саркоплазмой и содержащей большое количество калия, магния и фосфатов, а также множество ферментов. Кроме того, имеется огромное число митохондрий, расположенных параллельно миофибриллам. Они снабжают сокращающиеся миофибриллы большим количеством энергии в форме аденозинтрифосфата, синтезируемого митохондриями.
Саркоплазматический ретикулум. В саркоплазме, окружающей миофибриллы каждого мышечного волокна, имеется также хорошо развитый ретикулум, называемый саркоплазматическим ретикулумом. Он имеет специфическую организацию, исключительно важную для регуляции мышечного сокращения . Саркоплазматический ретикулум особенно сильно развит в очень быстро сокращающихся типах мышечных волокон.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
ПОПЕРЕ́ЧНО-ПОЛОСА́ТЫЕ МЫ́ШЦЫ
ПОПЕРЕ́ЧНО-ПОЛОСА́ТЫЕ МЫ́ШЦЫ (поперечно-полосатая мышечная ткань), сократимая ткань позвоночных животных и человека, формирующая скелетную (туловищную, или соматическую) мускулатуру, мимические мышцы, язык, часть стенки пищевода. Развивается из среднего зародышевого листка ( мезодер мы) и состоит из многоядерных мышечных волокон (симпластов), покрытых возбудимой плазматич. мембраной (сарколеммой), сходной по электрич. свойствам с мембранами нервных клеток. Под плазматич. мембраной мышечных волокон расположены также миосателлиты – мелкие одноядерные малодифференцированные клетки, лишённые сократимых белков и служащие для роста и регенерации П.-п. м. Группы волокон образуют мышечные пучки, которые, объединяясь, формируют мышцу. В соединит. ткани, окружающей мышечные волокна, мышечные пучки и всю мышцу, проходят кровеносные сосуды и нервы. Своими концами П.-п. м. прочно соединяются с сухожилием, с помощью которого прикрепляются обычно к костям или хрящам. Функциональной единицей мышечных волокон являются миофибриллы – тонкие нити, обеспечивающие мышечное сокращение . Для П.-п. м. характерна видимая в световой микроскоп поперечная исчерченность (отсюда назв.), обусловленная чередованием в миофибриллах участков с разными физико-химич. и оптич. свойствами.
Мышечная ткань: строение и функции
Ткань — сочетание похожих по строению клеток, выполняющих общие функции. Мышечная ткань в организме многоклеточного животного и человека отвечает за движения, механическую прочность и защиту внутренних органов. Ходьба, продвижение пищи, биение сердца — функции, выполняемые различными мышцами.
Строение и функции
Клеточные элементы мышечной ткани вытянуты в длину, за что получили название «волокна». Цитоплазма клеток содержит тонкие белковые нити миофибриллы, которые могут удлиняться и укорачиваться (табл. 1). Специальные органеллы, выработка энергии митохондриями обеспечивают сокращение и растяжение волокон.
Строение и функции мышечной ткани
Виды мышечной ткани
Расположение в организме
Поперечно-полосатая
Состоит из длинных и толстых волокон (рис. 1). Они образованы путем слияния отдельных клеток. Ядер много. Полосатая исчерченность вызвана чередованием светлых и темных дисков. Волокна объединяются в пучки.
Произвольные движения тела, дыхание, мимика лица и ряд других действий.
Основа скелетных мышц, языка, глотки, начальной части пищевода.
Отдельные веретеновидные клетки имеют небольшие размеры, объединены в пучки (по 5–10 шт.). В каждой клетке одно ядро (рис. 1). Тонкие миофибриллы протянулись между концами клетки. Ткань лишена поперечной полосатости.
Непроизвольные сокращения стенок внутренних органов с под влиянием нервных импульсов.
Мышечные слои кожи и внутренних органов (пищеварительной системы, мочевого пузыря, кровеносных и лимфатических сосудов, матки).
Поперечно-полосатая сердечная
Клетки удлиненные, разветвленной формы, с небольшим количеством ядер, образуют единую сеть (рис. 1). Поперечная полосатость возникает за счет блестящих полосок на соединениях между клетками.
«Двигатель» кровообращения. Непроизвольные сокращения сердечной мышцы могут происходить под управлением вегетативного отдела нервной системы.
Мышечные ткани обеспечивает передвижение организма в пространстве. Сокращения мышц необходимы для изменения положения отдельных частей тела. Мышцы, помимо двигательной, выполняют защитную и теплообменную функции.
Свойства
Мышечное волокно растягивается, но в состоянии покоя возвращается к своим первоначальным размерам. Это свойство — результат взаимодействия белковых нитей миофибрилл в цитоплазме клеток. Каждая миофибрилла состоит из протофибрилл: тонких, образованных актином, и более толстых — из миозина.
- электрическая возбудимость;
- сократимость;
- проводимость;
- растяжимость;
- эластичность.
Мышечная ткань способна к произвольным или непроизвольным сокращениям в ответ на нервные импульсы. Происходит взаимодействие фибриллярных белков — актина и миозина. В этом процессе обязательно участвуют неорганические ионы кальция. При сокращении тонкие нити актина скользят по толстым протофибриллам миозина.
Сравнительная характеристика видов мышечной ткани
В теле позвоночных животных и человека три типа мышечной ткани: поперечнополосатая, гладкая, сердечная. В организме низших животных мышцы состоят из гладкой ткани. У позвоночных животных и человека этот тип ткани образует стенки внутренних органов, кроме сердца (рис. 2).
Гладкая мышечная ткань
Медленные и продолжительные сокращения мышц контролирует вегетативная нервная система. Задача таких движений — сохранить или изменить объем полых органов против сил растяжения. Гладкие мышцы сокращаются и растягиваются больше, чем другие типы мышечной ткани. Сокращение длится намного дольше, что связано со скоростью прохождения ионов кальция, регулирующих процесс.
- сокращаются в 10–20 раз медленнее, чем скелетные;
- способны к длительным сокращениям;
- не затрачивают много энергии;
- медленнее наступает утомление.
Сокращения гладкой мышечной ткани происходят непроизвольно, то есть независимо от воли человека. Сигнал нервной системы проходит через всю массу клеток, что объясняется особенностями иннервации гладкой мускулатуры.
Поперечнополосатая ткань
Клетки имеют толщину от 10 до 100 мкм, длину от 10 до 40 см. Цитоплазма содержит большое количество ядер и миофибрилл, занимающих центральное положение (рис. 2). В зрелых клетках насчитывается сотни миофибрилл, более 100 ядер. Актиновые и миозиновые нити внутри миофибрилл сцеплены друг с другом (рис. 3). Способность к быстрому сокращению у этой ткани выше, чем у других.
Читайте также: Характеристики ткани для обуви
Мышечные волокна покрыты оболочкой — сарколеммой. Есть чередующиеся пластинки белков разной плотности, обладающие неодинаковыми коэффициентами преломления света. В оптический микроскоп такие мышцы кажутся исчерченными поперек. Сократительные элементы объединены в мышечные пучки, покрытые соединительнотканной оболочкой. Скелетные мышцы хорошо снабжены кровеносными сосудами и нервами.
Поперечнополосатая сердечная ткань
Особые свойства сердечной мышцы обусловлены строением волокон. Клетки длиной до 100 мкм встречаются только в сердце, не сливаются, как в поперечнополосатой мышечной ткани (рис. 2). Расположение актина и миозина, диски в мышце сердца такие же, как в волокнах скелетной мышечной ткани. Отличительная особенность — наличие глянцевых полосок в местах соединения клеток. Благодаря соединению волокон в единую сеть, возбуждение на одном участке быстро охватывает мышечную массу, участвующую в сокращении.
Мышечная ткань сердца способна к автоматической работе. Между сокращениями наступает рефракторный период, когда мышца находится в покое. При сокращении происходит уменьшении просвета полостей сердца — предсердий и желудочков.
Сердечная поперечнополосатая ткань сокращается в 10–15 раз дольше, чем скелетные мышцы. В нормальных условиях у человека сокращение и расслабление происходит 70–80 раз в минуту. Сокращение вызывают электрические импульсы, возникающие в самом сердце. Этот процесс связан носителем энергии — аденозинтрифосфатом (АТФ).
Полностью автономная работа, непрерывная ритмическая активность — физиологические отличия сердечной мышцы от скелетных. Нервные импульсы вегетативной нервной системы, иннервирующей сердце, не требуются для бесперебойной работы органа.
Поперечно полосатая мышечная ткань белки
1. Классификация и функции мышечной ткани.
2. Поперечно-полосатая мышечная ткань.
5. Гистогенез и регенерация мышечной ткани.
1. Классификация и функции мышечной ткани
Уникальная способность животных организмов к активному движению осуществляется благодаря мышечной ткани. Сократимость – это свойство только мышечной ткани в нашем организме. Кроме сократимости она обладает возбудимостью и проводимостью. Имея разное происхождение и строение, мышечные ткани объединяет способность к сокращению. Сократительный аппарат занимает значительную часть в цитоплазме, в его составе присутствуют актиновые и миозиновые филаменты из сократительных белков – актин и миозин, которые формируют органеллы специального значения–миофибриллы.
По морфофункциональному признаку различают:
· Скелетную или поперечно-полосатую или исчерченную мышечную ткань . Начало и прикрепление мышц находится на скелете. Мышцы являются произвольными, поскольку их сокращения подчиняются нашей воле. К этой группе мышц относят скелетные мышцы, мышцы языка, гортани и др.
· Сердечная мышечная ткань входит в состав мышечной стенки сердца. Иннервируется вегетативными нервами, является непроизвольной.
· Гладкая (неисчерченная) мышечная ткань характеризуется отсутствием исчерченности , а поскольку сокращения также не подчиняются нашей воле, то мышцы называют непроизвольными. Иннервация осуществляется вегетативной нервной системой. Из гладких мышц построены стенки внутренних органов, стенка сосудов.
В зависимости от источников развития выделяют пять типов мышечной ткани:
1. Мезенхимного происхождения (гладкая мышечная ткань).
2. Из кожной эктодермы и прехордальной пластинки – миоэпителиальные клетки (например, в потовых, слюнных железах).
3. Нейральное происхождение (из нервной трубки) – мышцы суживающие и расширяющие зрачок.
4. Целомическое происхождение ( миоэпикардиальная пластинка) – сердечная мышечная ткань.
5. Из миотомов мезодермы – исчерченная мышечная ткань.
1. Движение организма, его перемещение в пространстве.
2. Обеспечение физиологических процессов дыхания, кровообращения, пищеварения, внешней секреции.
3. Участие в теплорегуляции.
4. Запасающая (депо гликогена).
2. Поперечно-полосатая мышечная ткань
Источником развития являются клетки миотомов миобласты. Различают головные, шейные, грудные, поясничные, крестцовые миотомы . Они разрастаются в дорзальном и вентральном направлениях. В них рано врастают ветви спинномозговых нервов.
Часть миобластов дифференцируется на месте (образуют аутохтонную мускулатуру), а другие, с 3 недели внутриутробного развития мигрируют в мезенхиму и, сливаясь, друг с другом образуют мышечные трубки ( миотубы ) с крупными центрально ориентированными ядрами. В миотубах происходит дифференцировка специальных органелл миофибрилл. Первоначально они располагаются под плазмолеммой, а затем заполняют большую часть миотубы . Ядра смещаются к периферии. Клеточные центры и микротрубочки исчезают. Гранулярная ЭПС значительно редуцируется. Такая многоядерная структура называется симпласт , а для мышечной ткани — миосимпласт .
Часть миобластов дифференцируется в миосателлитоциты , которые располагаются на поверхности миосимпластов и принимают участие в регенерации мышечной ткани.
Структурной единицей мышечной ткани является мышечное волокно , состоящее из миосимпласта и миосателлитоцитов , покрытых общей базальной мембраной. Длина мышечных волокон колеблется от 1 до 40 мм, а толщина 0,1 мм.
В мышечном волокне различают мембранный аппарат, фибриллярный (сократительный) аппарат, трофический аппарат (ядро, саркоплазма, цитоплазматические органеллы).
Мембранный аппарат . Каждое мышечное волокно покрыто сарколеммой, где различают наружную базальную мембрану и плазмолемму (под базальной мембраной), плазмолемма образует впячивания (Т-трубочки).
К плазмолемме снаружи прилежат миосателлитоциты . При повреждении базальной мембраны запускается митотический цикл миосателлитоцитов .
Фибриллярный аппарат . Исчерченные волокна можно разделить на составляющие их фибриллы (диаметром 1 мкм), названные миофибриллами. В мышечном волокне они ориентированы продольно.
При рассматривании мышечных волокон и миофибрилл под световым микроскопом, отмечается чередование в них темных и светлых участков – дисков. Темные диски отличаются двойным лучепреломлением и называются анизотропными дисками или А- дисками. Светлые диски не обладают двойным лучепреломлением и называются изотропными или I – дисками. В средней части диска А имеется более светлый участок Н-зона (участок содержащий только толстые нити белка миозина). В области Н-зоны выделяется более темная М-линия, состоящая из миомезина (необходим для сборки толстых нитей и их фиксации при сокращении). В середине диска I расположена плотная линия Z, которая построена из белковых фибриллярных молекул. В частности, большую роль играет альфа актинин . Z – линия соединена с соседними миофибриллами с помощью белка десмина и поэтому все названные линии и диски соседних миофибрилл совпадают и создается картина поперечно-полосатой исчерченности мышечного волокна.
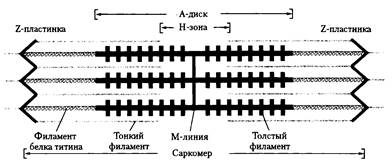
Структурной единицей миофибриллы является саркомер (S) – это пучок миофиламентов заключенный между двумя Z линиями ( рис.45 ). Принимая во внимание вышеуказанные обозначения можно структуру саркомера записать в виде формулы:
Под электронным микроскопом миофибриллы представляют агрегаты из толстых ( меозиновых ) филаментов (диаметр 14 нм , длина 1500 нм , расстояние между ними 20-30 нм ). Между толстыми филаментами располагаются тонкие филаменты ( диаметр 7-8 нм ).
Толстые филаменты ( миозиновые нити) состоят из молекул белка миозина. Он является важнейшим сократительным белком мышцы. При непосредственном участии миозина химическая энергия трансформируется в механическую работу. Каждая миозиновая нить состоит из 300-400 молекул миозина. Молекула миозина – это гексамер , состоящий из двух тяжелых и четырех легких цепей. Тяжелые цепи представляют собой две спирально закрученные полипептидные нити. Они несут на своих концах глобулярные (шаровидные) головки. Между головкой и тяжелой цепью – шарнирный участок, с помощью которого головка может изменять свою конфигурацию. В области головок — легкие цепи (по две на каждой). Молекулы миозина уложены в толстой нити таким образом, что их головки обращены наружу, выступая над поверхностью толстой нити, а тяжелые цепи образуют стержень толстой нити.
Тяжелые и легкие цепи в молекуле миозина можно разделить обработкой мочевиной, гуанидинхлоридом и др. При мягкой обработке можно отделить только легкие цепи. Миозину свойственна АТФ- азная активность – высвобождающаяся энергия используется для мышечного сокращения.
Тонкие нити ( актиновые нити). Образованы тремя белками: актином, тропонином и тропомиозином . Основным по массе белком является актин, который образует спираль. Молекулы тропомиозина располагаются в желобке этой спирали, молекулы тропонина располагаются вдоль спирали.
Толстые нити занимают центральную часть саркомера –А-диск, тонкие занимают I диски и частично входят между толстыми миофиламентами . Только толстые нити содержит Н-зона.
При поступлении нервных импульсов по аксонам двигательных нейронов происходит сокращение мышечного волокна. Каждое мышечное волокно имеет собственный аппарат иннервации (моторная бляшка) и окружено сетью гемокапилляров , располагающихся в прилежащей рыхлой соединительной ткани. Этот комплекс называется мион . Группа мышечных волокон, которые иннервируются одним мотонейроном называется нервно-мышечной единицей. Мышечные волокна в этом случае могут располагаться не рядом (одно нервное окончание может контролировать от одного до десятков мышечных волокон).
В покое взаимодействие тонких и толстых нитей ( миофиламентов ) невозможно, т.к. миозин-связывающие участки актина заблокированы тропомиозином . При высокой концентрац ии ио нов кальция конформационные (пространственные) изменения тропомиозина приводят к разблокированию миозин-связывающих участков молекул актина. Плазмолемма миосимпласта образует пальцевидные впячивания (инвагинации) ориентированные поперечно по отношению к миосимпласту называемые Т-трубочки. К каждой Т-трубочке примыкают по две цистерны саркоплазматического ретикулума (гладкая ЭПС), образуя триаду: две цистерны и одна Т-трубочка. Са 2 + концентрируется в цистернах (там его концентрация в 800 раз больше, чем в саркоплазме).
Механизм сокращения. При поступлении нервного импульса волна деполяризации доходит до цистерн саркоплазматического ретикулума , из них выделяются ионы кальция и концентрация кальция в саркоплазме резко возрастает. Са 2 + диффундирует к тонким нитям ( филаментам ) саркомера , где связывается с тропонином и миозиновыми головками. Это приводит:
1. К изменению конформации (пространственного расположения) тропомиозина , что, в свою очередь, приводит к освобождению участков актина, необходимых для взаимодействия с миозиновыми головками.
2. Появлению АТФ- азной активности миозина.
3. Взаимодействию миозиновых головок с актином ( актин о — миозиновые «мостики») и скольжению нитей (рис. 46).
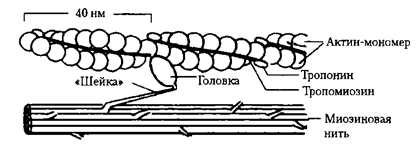
Рис. 46 . Актомиозиновый комплекс.
Все это вместе взятое приводит к тому, что миозиновые головки «шагают» по актину, образуя в ходе перемещения новые связи актина и миозина, сближая две Z-линии. При сокращении уменьшаются только светлые диски.
Читайте также: Как сделать из ткани сумочку для куклы
Расслабление. Са 2 + -АТФ-аза саркоплазматического ретикулума закачивает Са 2+ из саркоплазмы в цистерны. В саркоплазме концентрация Са 2 + становится низкой. Са 2 + — тропомиозин закрывает миозин-связывающие участки тонких нитей и препятствует их взаимодействию с миозином.
Чувствительная иннервация (нервно-мышечные веретена). Интрафузальные мышечные волокна вместе с чувствительными нервными окончаниями формируют нервно-мышечные веретена, являющиеся рецепторами скелетной мышцы. Снаружи сформирована капсула веретена. При сокращении поперечно-полосатых (исчерченных) мышечных волокон изменяется натяжение соединительно-тканной капсулы веретена и соответственно изменяется тонус интрафузальных (расположенных под капсулой) мышечных волокон. Формируется нервный импульс.
Классификация и типы мышечных волокон . Скелетные мышцы, состоящие из мышечных волокон отличаются по многим параметрам: цвету, диаметру, утомляемости, скорости сокращения и т.д. В каждой мышце присутствуют разные типы мышечных волокон. В исчерченных мышцах различают два вида мышечных волокон: экстрафузальные , которые преобладают и обуславливают собственно сократительную функцию мышцы и интрафузальные , входящие в состав проприоцепторов–нервно-мышечных веретен.
По характеру сокращения мышечные волокона делят на фазные и тонические. Фазные способны осуществлять быстрые сокращения, но не могут длительно удерживать достигнутый уровень укорочения. Тонические –о беспечивают поддержание статического напряжения или тонуса.
По биохимическим особенностям и цвету выделяют красные и белые мышечные волокна. Цвет мышцы обусловлен степенью васкуляризации . Кроме того, существует прямая корреляция между содержанием миоглобина и цветом мышцы. Характерной особенностью красных мышечных волокон является наличие многочисленных митохондрий, цепи которых располагаются между миофибриллами. В белых мышечных волокнах митохондрий меньше и они располагаются равномерно в саркоплазме мышечного волокна.
Скорость сокращения определяется типом миозина . Высокую скорость сокращения обеспечивает быстрый миозин (для него характерна высокая активность АТФ-азы); меньшая скорость сокращения характерна для медленного миозина (характерна невысокая активность АТФ-азы). Следовательно, по активности АТФ-азы можно судить и о наборе миозинов.
Тип окислительного обмена . Мышечные волокна используют два пути образования АТФ:
* при анаэробном типе метаболизма из 1 молекулы глюкозы образуется 2 молекулы АТФ и молочная кислота.
* при аэробном окислении из 1 молекулы глюкозы образуется 38 молекул АТФ и конечные продукты метаболизма: СО 2 и Н2О. Идентификация мышечных волокон основана на выявлении активности фермента сукцинатдегидрогеназы (СДГ), которая является маркером для митохондрий и цикла Кребса. Активность этого фермента свидетельствует о напряженности энергетического метаболизма. Выделяют мышечные волокна А-типа (гликолитические) с низкой активностью СДГ, С-тип ( оксидативные ) с высокой активностью СДГ. Мышечные волокна В-типа занимают промежуточное положение. Переход мышечных волоко от А-типа в С-тип маркирует изменения от анаеробного гликолиза к метаболизму, зависящему от кислорода.
Существует много и других классификаций.
Факторами, определяющими структуру и функцию скелетной мышцы являются влияние нервной ткани, гормональное влияние, уровень васкуляризации , уровень двигательной активности и местоположение мышцы.
3. Сердечная мышечная ткань
Она находится в мышечной оболочке сердца (миокард) и в устьях связанных с ним крупных сосудов. Имеет клеточный тип строения и основным функциональным свойством служит способность к спонтанным ритмическим сокращениям.
Развивается из миоэпикардиальной пластинки (висцеральный листок спланхнотома в шейном отделе), клетки которой размножаются митозом а потом дифференцируются. В клетках появляются миофиламенты , которые далее формируют миофибриллы.
Сердечная мышечная ткань образована клетками, которые называются кардиомиоциты . Между ними расположена рыхлая соединительная ткань и кровеносные сосуды с помощью вставочных дисков. Кардиомиоциты обьединяются в мышечные «волокна». Продольные и боковые связи кардиомиоцитов обеспечивают функциональное единство миокарда. Последние являются комплексом контактов. На поперечном срезе вставочных дисков выявляют десмосомы и щелевидные контакты ( нексусы ).
Выделяют рабочие (сократительные) кардиомиоциты , которые образуют цепочки клеток и обеспечивают силу сокращения сердечной мышцы. Клетки удлиненной формы с центрально расположенным ядром (рис.47). Вблизи ядра (или двух) комплекс Гольджи и гранулы гликогена. Между миофибриллами лежат многочисленные митохондрии. Имеются Т-трубочки и L-трубочки. Десмосомы обеспечивают механическое сцепление, которое препятствует расхождению кардиомиоцитов . Щелевидные контакты способствует передаче сокращения от одного кардиомиоцита к другому.
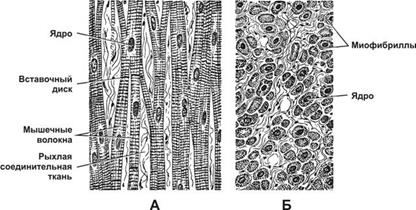
Рис. 47. Миокард (А – продольный, Б – поперечный разрез).
Проводящие (атипичные) кардиомиоциты – среди них различают:
1. Водители ритма – это клетки небольших размеров, в саркоплазме мало гликогена, мало миофибрилл и они расположены по периферии. Клетки имеют хорошее кровоснабжение и иннервацию. Они воспринимают сигналы от нервных окончаний и способны автоматически генерировать сигналы обеспечивающие ритмические сокращения сердца.
2. Проводящие (переходные) кардиомиоциты проводят возбуждение от водителя ритма. Образуют длинные волокна. Миофибриллы в небольшом количестве, имеют спиральный ход, мелкие митохондрии, немного гликогена.
3. Волокна Пуркинье – являются самыми крупными клетками в мышечной ткани сердца с неупорядоченным расположением миофибрилл, множеством мелких митохондрий, много гликогена, нет Т-трубочек, клетки связаны между собой десмосомами и щелевидными контактами.
Секреторные кардиомиоциты – находятся в, основном, в предсердиях, преимущественно в правом. Характеризуются отростчатой формой и слабым развитием сократительного аппарата. В саркоплазме, вблизи полюсов ядра-секреторные гранулы, содержащие атриопептин (гормон, регулирующий артериальное давление). Гормон вызывает потерю натрия и воды с мочой, расширение сосудов, снижение давления, угнетение секреции альдостерона, кортизола, вазопрессина.
Сократительный аппарат рабочих кардиомиоцитов сходен со скелетными мышечными волокнами. Миофибриллы в кардиомиоците могут объединяться в комплексы, образуя единые сократительные структуры. В саркоплазме миофибриллы ориентированы продольно и располагаются преимущественно по периферии. Саркотубулярная система в целом имеет сходство с исчерченными мышечными волокнами. Саркоплазматическая сеть развита слабее, не так активно накапливает Са 2 . . При расслаблении, ионы кальция выделяются в саркоплазму с низкой скоростью, что обеспечивает автоматизм и частые сокращения кардиомиоцитов . Т-трубочки широкие и образуют диады (одна Т-трубочка и одна цистерна сети), которые сходятся в области Z-линии. Энергетический аппарат-это митохондрии и включения.
Структурно-функциональной единицей данной ткани являются гладкие мышечные клетки (ГМК), которые способны к гипертрофии и регенерации. Они образуют стенки внутренних полых органов, сосудов. Более крупные по размерам ГМК характерны для стенок полых внутренних органов, а меньших размеров – для стенки сосудов. Клетки контролируют моторику, величину просвета. Имеют веретенообразную форму, в центре палочковидной формы ядро. В ГМК отсутствует поперечно-полосатая исчерченность . ГМК окружены сарколеммой, которая снаружи покрыта базальной мембраной. Длина от 20 мкм до 1 мм. В саркоплазме у полюсов находится комплекс Гольджи , много митохондрий, рибосом, развит саркоплазматический ретикулум .
Миофиламенты расположены вдоль продольной оси. В ГМК актиновые и миозиновые филаменты не формируют миофибрилл. Актиновые нити (тонкие филаменты ) ориентированы по продольной оси ГМК. По количеству их больше и они прикрепляются к плотным тельцам, которые являются специальными сшивающими белками. Рядом с актиновыми нитями располагаются мономеры миозина ( микромиозин ). Обладая разной длиной они, значительно короче тонких нитей.
Сокращение гладких мышечных клеток осуществляется при взаимодействии актиновых филаментов и миозина. Сигнал идущий по нервным волокнам обуславливает выделение медиатора, что изменяет состояние сарколеммы. Она образует колбовидные впячивания ( кавеолы ), где концентрируются ионы кальция. Сокращение ГМК индуцируется притоком ионов кальция в саркоплазму ( кавеолы отшнуровываются и вместе с ионами кальция попадают в саркоплазму). Это приводит к полимеризации миозина и взаимодействию его с актином. Актиновые нити и плотные тельца сближаются, усилие передается на сарколемму и ГМК укорачивается. Миозин ГМК способен взаимодействовать с актином только после фосфорилирования его легких цепей особым ферментом– киназой легких цепей. После прекращения сигнала ионы кальция покидают кавеолы ; миозин деполяризуется, теряет сродство к актину. В результате комплексы миофиламентов распадаются; сокращение прекращается.
Особые типы гладких мышечных клеток . Миоэпителиальные клетки являются производными эктодермы, не имеют исчерченности . Окружают секреторные отделы и выводные протоки желез (слюнных, молочных, слезных). С железистыми клетками они связаны десмосомами. Сокращаясь, способствуют выделению секрета. В концевых (секреторных) отделах форма клеток отросчатая , звездчатая. Ядро в центре, в цитоплазме, преимущественно в отростках локализованы миофиламенты , которые образуют сократительный аппарат. В этих клетках есть и цитокератиновые промежуточные филаменты , что подчеркивает их сходство с эпителиоцитами .
Мионейральные клетки – развиваются из клеток наружного слоя глазного бокала и образуют мышцу, суживающую зрачок и мышцу, расширяющую зрачок. По строению первая мышца сходна с ГМК мезенхимного происхождения. Мышца, расширяющая зрачок образована отростками клеток, располагающимися радиально, а ядросодержащая часть клетки находится между пигментным эпителием и стромой радужки.
Миофибробласты относятся к рыхлой соединительной ткани и представляют собой видоизмененные фибробласты. Они проявляют свойства как фибробластов, так и ГМК (обладают выраженными сократительными свойствами). Как вариант этих клеток можно рассматривать миоидные клетки в составе стенки извитого семенного канальца яичка и наружного слоя теки фолликула яичника. При заживлении раны часть фибробластов синтезирует гладкомышечные актины и миозины.
Эндокринные гладкие миоциты – это видоизмененные ГМК, представляющие основной компонент юкста-гломерулярного аппарата почек. Они находятся в стенке артериол почечного тельца, имеют хорошо развитый синтетический аппарат и редуцированный сократительный. Продуцируют фермент ренин, находящийся в гранулах и попадающий в кровь механизмом экзоцитоза .
5. Гистогенез и регенерация мышечной ткани
Гистогенез скелетной мышечной ткани . Источником развития элементов скелетной (соматической) поперечнополосатой мышечной ткани являются клетки миотомов мезодермы – миобласты. Одни из них дифференцируются на месте и участвуют в образовании так называемых аутохтонных мышц. Другие клетки мигрируют из миотомов в мезенхиму. Они уже детерминированы, хотя внешне не отличаются от других клеток мезенхимы. Их дифференцировка продолжается в местах закладки других мышц тела. В ходе дифференцировки возникают две клеточные линии. Клетки одной из линий сливаются, образуя удлиненные симпласты – мышечные трубочки ( миотубы ). В них происходит дифференцировка специальных органелл – миофибрилл. В это время в миотубах отмечается хорошо развитая гранулярная эндоплазматическая сеть. Миофибриллы сначала располагаются под плазмолеммой, а затем заполняют большую часть миотубы . Ядра, напротив, из центральных отделов смещаются к периферии. Клеточные центры и микротрубочки при этом полностью исчезают. Гранулярная эндоплазматическая сеть редуцируется в значительной степени. Такие дефинитивные структуры называют миосимпластами . Клетки другой линии остаются самостоятельными и дифференцируются в миосателлитоциты ( миосателлиты ). Эти клетки располагаются на поверхности миосимпластов .
Читайте также: Зародышевая соединительная ткань человека
Значит, из миотомов мезодермы в определенные участки мезенхимы выселяются малодифференцированные клетки — миобласты, часть из которых выстраивается в виде цепочки в стык друг к другу и в области контактов миобластов цитолеммы исчезает и образуется симпластическое образование — миотрубка , в которой ядра в виде цепочки располагаются в середине, а по периферии начинают дифференцироваться из миофиламентов миофибриллы. К миотрубке подрастают нервные волокна, образуя двигательные нервные окончания. Под влиянием эфферентной нервной импульсации начинается перестройка мышечной трубки в мышечное волокно: ядра перемещаются на периферию симпласта к плазмолемме, а миофибриллы занимают его центральную часть, из гладкой эндоплазматической сети развивается саркоплазматическая сеть, окружающая каждую миофибриллу на всем ее протяжении. Плазмолемма миосимпласта образует глубокие трубчатые впячивания — Т-канальцы. За счет деятельности зернистой эндоплазматической сети вначале миобластов, а затем и мышечных труб синтезируются и выделяются с помощью пластинчатого комплекса белки и полисахариды, из которых формируется базальная пластинка мышечного волокна.
Следует отметить, что при формировании миотрубки , а затем и дифференцировки мышечного волокна часть миобластов не входит в состав симпласта , а прилежит к нему, располагаясь под базальной пластинкой. Эти клетки носят название миосателлитов и играют важную роль в процессах физиологической и репаративной регенерации. Установлено, что закладка поперечно-полосатых скелетных мышечных волокон ( миогенез ) происходит только в эмбриональный период. В постнатальном периоде осуществляется их дальнейшая дифференцировка и гипертрофия, но количество мышечных волокон даже в условиях интенсивной тренировки не увеличивается.
Гистогенез сердечной мышечной ткани . Источники развития сердечной поперечнополосатой мышечной ткани – симметричные участки висцерального листка спланхнотома в шейной части зародыша – миоэпикардиальные пластинки. Из них дифференцируются также клетки мезотелия эпикарда. В ходе гистогенеза стволовые клетки сердечной мышечной ткани (в составе миоэпикардиальной пластинки) в процессе дифференцировки дают начало нескольким клеточным дифферонам : возникает 5 видов кардиомиоцитов – рабочие (сократительные), синусные ( ритмзадающие или пейсмекерные ), переходные, проводящие, а также секреторные.
Исходными клетками сердечной мышечной ткани являются кардиомиобласты – уплощенные клетки (крупное ядро, светлая цитоплазма с небольшим количество рибосом и митохондрий). Происходит развитие комплекса Гольджи , гранулярной ЭПС. Клетки обладают высокой способностью к делению. Кардиомиобласты дифференцируются в кардиомиоциты . В цитоплазме кардиомиоцитов увеличивается число полисом, канальцев гранулярной ЭПС, накапливаются гранулы гликогена, увеличивается объем актомиозинового комплекса. Кардиомиоциты путем контактов «конец в конец», «конец в бок» формируют клеточные комплексы – сердечные мышечные волокна.
Гистогенез гладкой мышечной ткани . Этот вид мышечной ткани расположен в стенках органов и имеет генез мезенхимного типа. Миоциты объединяются в пучки, между которыми располагаются тонкие прослойки соединительной ткани. В эти прослойки вплетаются ретикулярные и эластические волокна, окружающие миоциты . В прослойках проходят кровеносные сосуды и нервные волокна. Терминали последних оканчиваются не непосредственно на миоцитах , а между ними. Поэтому после поступления нервного импульса медиатор распространяется диффузно, возбуждая сразу многие клетки. Гладкая мышечная ткань мезенхимного происхождения представлена главным образом в стенках кровеносных сосудов и многих трубчатых внутренних органов, а также образует отдельные мелкие мышцы (цилиарные). Гладкая мышечная ткань в составе конкретных органов имеет неодинаковые функциональные свойства. Это обусловлено тем, что на поверхности органов имеются разные рецепторы к конкретным биологически активным веществам. Поэтому и на многие лекарственные препараты их реакция неодинакова. Возможно, разные функциональные свойства тканей связаны и с конкретной молекулярной организацией актиновых филаментов .
Как было сказано выше, гладкие мышцы развиваются из мезенхимы. Клетки мезенхимы начинают активно размножаться и образуют сгущения. Первым признаков начавшейся дифференциации служит возникновение в цитоплазме клеток протофибрилл . Малодифференцированные клетки располагаются рыхло, соединяются между собой отростками (как клетки мезенхимы). Миофиламенты не имеют правильной ориентации. Позднее гладкомышечные клетки становятся веретенообразными и плотно прилегают друг к другу. Количество протофибрилл в них увеличивается, и они располагаются только в направлении длинной оси клетки.
В активированных миоцитах увеличивается синтез сократительных белков, происходит укрупнение и гиперхроматоз ядра, гипертрофия ядрышка, увеличивается количество свободных рибосом и полисом, наблюдается активация ферментов. Клеточная регенерация осуществляется как за счет активации миоцитов малого объема (камбиальных элементов), так и за счет дифференцированных клеток (обладающих способностью к митозу).
При функциональной нагрузке (беременности) или при патологии наблюдается увеличение массы гладкомышечной ткани за счет гипертрофии (увеличения размеров) или за счет гиперплазии (увеличения количества) гладких миоцитов
Мышечная ткань эпидермального происхождения Миоэпителиальные клетки развиваются из эпидермального зачатка. Они встречаются в потовых, молочных, слюнных и слезных железах и имеют общих предшественников с их секреторными клетками. Миоэпителиальные клетки непосредственно прилежат к собственно эпителиальным и имеют общую с ними базальную мембрану. При регенерации те и другие клетки тоже восстанавливаются из общих малодифференцированных предшественников. Большинство миоэпителиальных клеток имеют звездчатую форму. Эти клетки нередко называют корзинчатыми : их отростки охватывают концевые отделы и мелкие протоки желез. В теле клетки располагаются ядро и органеллы общего значения, а в отростках – сократительный аппарат, организованный, как и в клетках мышечной ткани мезенхимного типа.
Мышечная ткань нейрального происхождения. Миоциты этой ткани развиваются из клеток нейрального зачатка в составе внутренней стенки глазного бокала. Тела этих клеток располагаются в эпителии задней поверхности радужки. Каждая из них имеет отросток, который направляется в толщу радужки и ложится параллельно ее поверхности. В отростке находится сократительный аппарат, организованный так же, как и во всех гладких миоцитах . В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы – суживающую и расширяющую зрачок.
Регенерация. В мышечной, как в других тканях, различают два вида регенерации — физиологическую и репаративную . Из всех видов мышечной ткани лучше всего регенерирует гладкая, так как ее клетки миоциты могут активно делиться митозом всю жизнь.
Физиологическая регенерация скелетной мышечной ткани проявляется в форме гипертрофии мышечных волокон, что выражается в увеличении их толщины и даже длины, увеличение числа органелл, главным образом миофибрилл, а также нарастании числа ядер, что, в конечном счете, проявляется увеличением функциональной способности мышечного волокна. Радиоизотопным методом установлено, что увеличение числа ядер в мышечных волокнах в условиях гипертрофии достигается за счет деления клеток миосателлитов и последующего вхождения в миосимпласт дочерних клеток.
Увеличение числа миофибрилл осуществляется посредством синтеза актиновых и миозиновых белков свободными рибосомами и последующей сборки этих белков в актиновые и миозиновые миофиламенты параллельно с соответствующими филаментами саркомеров . В результате этого вначале происходит утолщение миофибрилл, а затем их расщепление и образование дочерних миофибрилл. Кроме того возможно образование новых актиновых и миозиновых миофиламентов не параллельно, а встык предшествующим миофибриллам, чем достигается их удлинение. Саркоплазматическая сеть и Т-канальцы в гипертрофирующемся волокне образуются за счет разрастания предшествующих элементов.
При определенных видах мышечной тренировки может формироваться преимущественно красный тип мышечных волокон (у стайеров) или белый тип мышечных волокон (у спринтеров). Возрастная гипертрофия мышечных волокон интенсивно проявляется с началом двигательной активности организма (1-2 года), что обусловлено, прежде всего усилением нервной стимуляции. В старческом возрасте, а также в условиях малой мышечной нагрузки наступает атрофия специальных и общих органелл, истончение мышечных волокон и снижение их функциональной способности.
Репаративная регенерация развивается после повреждения мышечных волокон. При этом способ регенерации зависит от величины дефекта. При значительных повреждениях на протяжении мышечного волокна миосателлиты в области повреждения и в прилежащих участках растормаживаются, усиленно пролиферируют, а затем мигрируют в область дефекта мышечного волокна, где выстраиваются в цепочки, формируя миотрубку . Последующая дифференцировка миотрубки приводит к восполнению дефекта и восстановлению целостности мышечного волокна. В условиях небольшого дефекта мышечного волокна на его концах, за счет регенерации внутриклеточных органелл, образуются мышечные почки, которые растут навстречу друг другу, а затем сливаются, приводя к закрытию дефекта. Однако , репаративная регенерация и восстановление целостности мышечных волокон могут осуществляться при определенных условиях: во-первых, при сохраненной двигательной иннервации мышечных волокон, во-вторых, если в область повреждения не попадают элементы соединительной ткани (фибробласты). Иначе на месте дефекта мышечного волокна развивается соединительно-тканный рубец.
Советским ученым А. Н. Студитским доказана возможность аутотрансплантации скелетной мышечной ткани и даже целых мышц при соблюдении определенных условий:
- механическое измельчение мышечной ткани трансплантата, с целью растормаживания клеток-сателлитов и последующей их пролиферации;
- помещение измельченной ткани в фасциальное ложе;
- подшивание двигательного нервного волокна к измельченному трансплантату;
- наличие сократительных движений мышц-антагонистов и синергистов.
Регенерация сердечной мышечной ткани. В сердечной мышечной ткани отсутствуют камбиальные клетки, а сами кардиомиоциты утрачивают способность делиться к моменту рождения ребенка или в первые месяцы жизни. Поэтому при регенерации ткани не образуются новые кардиомиоциты и волокна. Наблюдается гипертрофия (увеличение объема) сохранившихся клеток.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом