
Мышечные ткани — это ткани, для которых способность к сокращению является главным свойством. Мышечные ткани составляют активную часть опорно-двигательного аппарата (пассивной частью являются кости, соединения костей).
Общими свойствами всех мышечных тканей является сократимость и возбудимость. К данной группе тканей относятся гладкая, поперечнополосатая скелетная и поперечнополосатая сердечная мышечные ткани. Клетки мышечной ткани имеют хорошо развитый цитоскелет, содержат много митохондрий.

Гладкая (висцеральная) мускулатура
Эта мышечная ткань встречается в стенках внутренних органах (бронхи, кишечник, желудок, мочевой пузырь), в стенках сосудов, протоках желез. Эволюционно является наиболее древним видом мускулатуры.
Состоит из веретенообразных миоцитов — коротких одноядерных клеток. Между клетками имеются межклеточные контакты — нексусы (лат. nexus — связь). Благодаря нексусам возбуждение, возникшее в одной клетке, волнообразно распространяется на все остальные клетки.

Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы внутренних органов (к примеру мочевого пузыря), сокращается медленно, практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает — сокращается и утомляется быстро.
Осуществляется сокращение с помощью клеточных органоидов — миофиламентов, которые расположены в клетке хаотично и не имеют такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их тоже изучим).
Особо заметим, что в гладкой мышечной ткани миофиламенты собираются в миофибриллы только во время сокращения. У таких временных миофибрилл не может быть регулярной организации, а значит ни у таких миофибрилл, ни у гладких миоцитов не может быть поперечной исчерченности.
Гладкая мышечная ткань сокращается непроизвольно (неподвластна воле человека). Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой. К примеру невозможно по желанию сузить или расширить бронхи, кровеносные сосуды, зрачок.

Гладкая мышечная ткань называется неисчерченной, так как не обладает поперечной исчерченностью, характерной для поперечнополосатых скелетной и сердечной мышечных тканей.
Скелетная (поперечнополосатая) мышечная ткань
Скелетная мышечная ткань образует диафрагму (дыхательную мышцу), мускулатуру туловища, конечностей, головы, голосовых связок.
В отличие от гладкой мускулатуры, скелетная образована не отдельными одноядерными клетками, а длинными многоядерными волокнами, имеющими до 100 и более ядер — миосимпластами. Миосимпласт (греч. sim — вместе + plast — образованный) представляет совокупность слившихся клеток, имеет длину от нескольких миллиметров до нескольких сантиметров (соответствует длине мышцы).
Внутри миосимпласта находится саркоплазма, снаружи миосимпласт покрыт сарколеммой. Сократительные элементы — миофибриллы (лат. fibra — волоконце) — длинные тяжеобразные органеллы в миосимпласте (около 1400).

Характерная черта данной ткани — поперечная исчерченность, выражающаяся в равномерном чередовании светлых и темных полос на мышечном волокне. Это происходит потому, что границы саркомеров в соседних миофибриллах совпадают, вследствие чего все волокно приобретает поперечную исчерченность. Теперь самое время изучить микроскопическую основу мышцы — саркомер.
Саркомер (от греч. sarco — мясо (мышца) + mere — маленький)
Саркомер — элементарная сократительная единица поперечнополосатых мышц, структурная единица миофибриллы. В состав саркомера (и миофибриллы в целом) входят миофиламенты (лат. filamentum — нить) двух типов, которые обеспечивают сократимость мышечной ткани.
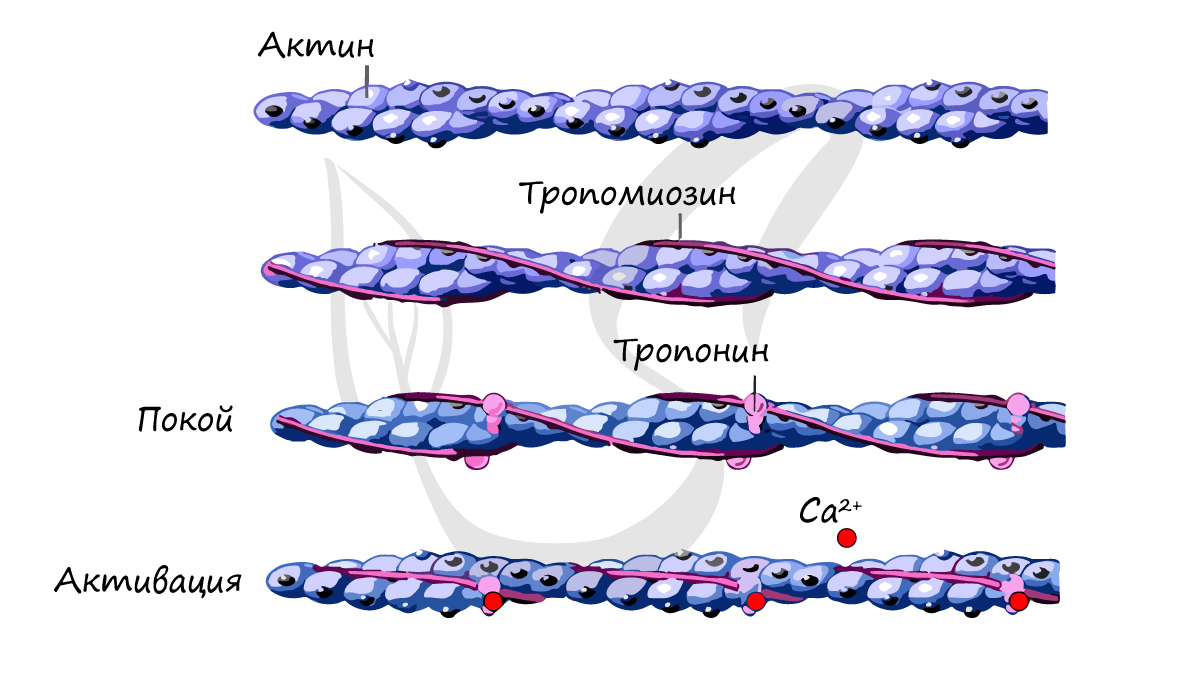
Саркомер состоит из актиновых (тонких) и миозиновых (толстых) филаментов, которые образованы главным образом белками актином и миозином. Сокращение происходит за счет взаимного перемещения миофиламентов: они тянутся навстречу друг другу, саркомер укорачивается (и мышца в целом).

Источником энергии для сокращения служат молекулы АТФ. К тому же невозможно представить сокращение мышц без участия ионов кальция: именно они связываются с тропонином, что приводит к изменению конформации тропомиозина (тропонин и тропомиозин — регуляторные белки между нитями актина), за счет чего становится возможно соединение актина и миозина. При сокращении мышц выделяется тепло (сократительный термогенез).
Замечу, что трупное окоченение (лат. rigor mortis) — посмертное затвердевание мышц — связано именно с ионами кальция, которые устремляются в область низкой концентрации (в саркоплазму миосимпласта), способствуя связыванию актина и миозина.
После смерти в мышце перестает синтезироваться АТФ, ее уровень быстро снижается. Как следствие этого перестает функционировать Ca-АТФаза — насос, выкачивающий ионы Ca из саркоплазмы в саркоплазматический ретикулум (мембранная органелла мышечных клеток (сходная с ЭПС), в которой запасаются ионы Ca).
Читайте также: Изготовление ткани из волокон хлопка
В саркоплазме повышается концентрация ионов Ca — замыкаются мостики между актином и миозином, однако разомкнуться они уже не могут, в связи с чем наблюдается стойкая мышечная контрактура (лат. contractura — стягивание, сужение): конечности очень сложно разогнуть или согнуть.

Вернемся к скелетным мышцам. Имеется еще ряд важных моментов, о которых нужно знать.
В процесс возбуждения вовлекается изолированно один миосимпласт, соседние миосимпласты (волокна) не возбуждают друг друга, в отличие от гладких миоцитов, где возбуждение предается между соседними клетками через нексусы. Скелетные мышцы сокращаются быстро и быстро утомляются (у гладких мышц фазы сокращения и расслабления растянуты во времени, мало утомляются) .
Скелетные мышцы сокращаются произвольно: они подконтрольны нашему сознанию. К примеру, по желанию мы можем изменить скорость движения руки, темп бега, силу прыжка. Мышцы покрыты фасцией, крепятся к костям сухожилиями, и, сокращаясь, приводят в движение суставы.

Сердечная поперечнополосатая мышечная ткань
Сердечная мышечная ткань образует мышечную оболочку сердца — миокард (от др.-греч. μῦς «мышца» + καρδία — «сердце»). Миокард — средний слой сердца, составляющий основную часть его массы. При работе сердечная мышечная ткань не утомляется.

Сердечная мышечная ткань состоит из кардиомиоцитов — одиночных клеток, имеющих поперечную исчерченность. Соединяясь друг с другом, кардиомиоциты образуют функциональные волокна.
Этот тип мышечной ткани удивительным образом сочетает свойства двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство — автоматизм.
Автоматизм — способность сердечной мышечной ткани возбуждаться и сокращаться самопроизвольно, без влияний извне. Это легко можно подтвердить, наблюдая сокращения изолированного сердца лягушки в физиологическом растворе: сокращения сердца в нем будут продолжаться несколько десятков минут после отделения сердца от организма.

Места контактов соседних кардиомиоцитов — вставочные диски (в их составе находятся нексусы), благодаря которым возбуждение одной клетки передается на соседние, таким образом волнообразно охватываются возбуждением и сокращаются новые участки миокарда.
Большое число контактов между кардиомиоцитами обеспечивает высокую эффективность и надежность проведения возбуждения по миокарду. Сокращается эта ткань непроизвольно, не утомляется.
На рисунке или микропрепарате узнать данную ткань можно по центральному положению ядер в клетках, поперечной исчерченности, наличию вставочных дисков и анастомозов (греч. anastomosis — отверстие) — мест соединений боковых поверхностей функциональных волокон (кардиомиоцитов).

В норме возбуждение проводится по проводящей системе сердца от предсердий к желудочкам (однонаправленно). Участок сердечной мышцы, в котором генерируются импульсы, определяющие частоту сердечных сокращений — водитель сердечного ритма.
Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных (англ. pacemaker — задающий ритм) клеток, которые также называют водителями ритма. Они спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.
Ответ мышц на физическую нагрузку
Физические нагрузки приводят к гипертрофии мышц (от др.-греч. ὑπερ- чрез, слишком + τροφή — еда, пища) — в них увеличивается количество мышечных волокон, объем мышечной массы нарастает.

В условиях гиподинамии (от греч. ὑπό — под и δύνᾰμις — сила), то есть пониженной активности, мышцы уменьшаются вплоть до полной атрофии (греч. а – «не» + trophe – питание). В худшем случае волокна мышечной ткани перерождаются в соединительную ткань, после чего пациент становится обездвиженным.

Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление. Гипертрофия сердца — состояние, требующее вмешательства врача и наблюдения за пациентом.
В большинстве случае гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы).

Происхождение мышц
Мышцы развиваются из среднего зародышевого листка — мезодермы.

© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
35. Мышечные ткани
Особенности строения: сократительный аппарат, занимающий значительную часть в цитоплазме структурных элементов мышечной ткани и состоящий из актиновых и миозиновых филаментов, которые формируют органеллы специального назначения –миофибриллы.
Читайте также: Изделия из ткани атлас
Классификация мышечных тканей
1. Морфофункциональная классификация:
1) Поперечнополосатая, или исчерченная мышечная ткань:скелетная и сердечная;
2) Неисчерченная мышечная ткань: гладкая.
2. Гистогенетическая классификация (в зависимости от источников развития):
1) Соматического типа (из миотомов сомитов) – скелетная мышечная ткань (поперечнополосатая);
2) Целомического типа (из миоэпикардиальной пластинки висцерального листка спланхнотома) – сердечная мышечная ткань (поперечнополосатая);
3) Мезенхимного типа (развивается из мезенхимы) – гладкая мышечная ткань;
4) Из кожной эктодермы и прехордальной пластинки – миоэпителиальные клетки желёз (гладкие миоциты);
5) Нейрального происхождения (из нервной трубки) – мионейральные клетки (гладкие мышцы, суживающие и расширяющие зрачок).
Функции мышечной ткани: перемещение тела или его частей в пространстве.
Исчерченная (поперечно-полосатая) мышечная ткань составляет до 40% массы взрослого человека, входит в состав скелетных мышц, мышц языка, гортани и др. Относятся к произвольным мышцам, поскольку их сокращения подчиняются воле человека. Именно эти мышцы задействованы при занятии спортом.
Гистогенез. Скелетная мышечная ткань развивается из клеток миотомов миобластов. Различают головные, шейные, грудные, поясничные, крестцовые миотомы. Они разрастаются в дорзальном и вентральном направлениях. В них рано врастают ветви спинномозговых нервов. Часть миобластов дифференцируется на месте (образуют аутохтонную мускулатуру), а другие с 3 недели внутриутробного развития мигрируют в мезенхиму и, сливаясь друг с другом, образуют мышечные трубки (миотубы) с крупными центрально ориентированными ядрами. В миотубах происходит дифференцировка специальных органелл миофибрилл. Первоначально они располагаются под плазмолеммой, а затем заполняют большую часть миотубы. Ядра смещаются к периферии. Клеточные центры и микротрубочки исчезают, грЭПС значительно редуцируется. Такая многоядерная структура называется симпласт, а для мышечной ткани – миосимпласт. Часть миобластов дифференцируется в миосателлитоциты, которые располагаются на поверхности миосимпластов и впоследствии принимают участие в регенерации мышечной ткани.
Строение скелетной мышечной ткани
Рассмотрим строение мышечной ткани на нескольких уровнях организации живого: на органном уровне (мышца как орган), на тканевом (непосредственно мышечная ткань), на клеточном (строение мышечного волокна), на субклеточном (строение миофибриллы) и на молекулярном уровне (строение актиновых и миозиновых нитей).
1 — мышца икроножная (органный уровень), 2 — поперечный срез мышцы (тканевой уровень) — мышечные волокна, между которыми РВСТ: 3 — эндомизий, 4 — нервное волокно, 5 — кровеносный сосуд; 6 — поперечный срез мышечного волокна (клеточный уровень): 7 — ядра мышечного волокна — симпласта, 8 — митохондрия между миофибриллами, синим цветом — саркоплазматический ретикулум; 9 — поперечный срез миофибриллы (субклеточный уровень): 10 — тонкие актиновые нити, 11 — толстые миозиновые нити, 12 — головки толстых миозиновых нитей.
1) Органный уровень: строение мышцы как органа.
Скелетная мышца состоит из пучков мышечных волокон, связанных воедино системой соединительнотканных компонентов. Эндомизий – прослойки РВСТ между мышечными волокнами, где проходят кровеносные сосуды, нервные окончания. Перимизий – окружает 10-100 пучков мышечных волокон. Эпимизий – наружная оболочка мышцы, представлена плотной волокнистой тканью.
2) Тканевой уровень: строение мышечной ткани.
Структурно-функциональной единицей скелетной поперечнополосатой (исчерченной) мышечной ткани является мышечное волокно – цилиндрической формы образование диаметром 50 мкм и длиной от 1 до 10-20 см. Мышечное волокно состоит из 1)миосимпласта (образование его смотри выше, строение – ниже), 2) мелких камбиальных клеток – миосателлитоцитов, прилежащих к поверхности миосимпласта и располагающиеся в углублениях его плазмолеммы, 3) базальной мембраны, которой покрыта плазмолемма. Комплекс плазмолеммы и базальной мембраны называется сарколемма. Для мышечного волокна характерна поперечная исчерченность, ядра смещены на периферию. Между мышечными волокнами – прослойки РВСТ (эндомизий).
3) Клеточный уровень: строение мышечного волокна (миосимпласта).
Термин «мышечное волокно» подразумевает «миосимпласт», поскольку миосимпласт обеспечивает функцию сокращения, миосателлитоциты участвуют только в регенерации.
Миосимпласт, как и клетка, состоит из 3-х компонентов: ядра (точнее множества ядер), цитоплазмы (саркоплазма) и плазмолеммы (которая покрыта базальной мембраной и называется сарколемма). Почти весь объём цитоплазмы заполнен миофибриллами – органеллами специального назначения, органеллы общего назначения: грЭПС, аЭПС, митохондрии, комплекс Гольджи, лизосомы, а также ядра смещены на периферию волокна.
В мышечном волокне (миосимпласте) различают функциональные аппараты: мембранный,фибриллярный (сократительный) и трофический.
Читайте также: Опухоли костной ткани мрт
Трофический аппарат включает ядра, саркоплазму и цитоплазматические органеллы: митохондрии (синтез энергии), грЭПС и комплекс Гольджи (синтез белков – структурных компонентов миофибрилл), лизосомы (фагоцитоз изношенных структурных компонентов волокна).
Мембранный аппарат: каждое мышечное волокно покрыто сарколеммой, где различают наружную базальную мембрану и плазмолемму (под базальной мембраной), которая образует впячивания (Т-трубочки). К каждой Т-трубочке примыкают по две цистернысаркоплазматического ретикулума (видоизменённая аЭПС), образуя триаду: две L-трубочки (цистерны аЭПС) и одна Т-трубочка (впячивание плазмолеммы). В цистернах аЭПС концентрируются Са 2+ , необходимый при сокращении. К плазмолемме снаружи прилежат миосателлитоциты. При повреждении базальной мембраны запускается митотический цикл миосателлитоцитов.
Фибриллярный аппарат. Большую часть цитоплазмы исчерченных волокон занимают органеллы специального назначения – миофибриллы, ориентированы продольно, обеспечивающие сократительную функцию ткани.
4) Субклеточный уровень: строение миофибриллы.
При исследовании мышечных волокон и миофибрилл под световым микроскопом, отмечается чередование в них темных и светлых участков – дисков. Темные диски отличаются двойным лучепреломлением и называются анизотропными дисками, или А— дисками. Светлые диски не обладают двойным лучепреломлением и называются изотропными, или I-дисками.
В середине диска А имеется более светлый участок – Н-зона, где содержатся только толстые нити белка миозина. В середине Н-зоны (значит и А-диска) выделяется более темная М-линия, состоящая из миомезина (необходим для сборки толстых нитей и их фиксации при сокращении). В середине диска I расположена плотная линия Z, которая построена из белковых фибриллярных молекул. Z-линия соединена с соседними миофибриллами с помощью белка десмина, и поэтому все названные линии и диски соседних миофибрилл совпадают и создается картина поперечнополосатой исчерченности мышечного волокна.
Структурной единицей миофибриллы является саркомер (S) — это пучок миофиламентов заключенный между двумя Z-линиями. Миофибрилла состоит из множества саркомеров. Формула, описывающая структуру саркомера:
5) Молекулярный уровень: строение актиновых и миозиновых филаментов.
Под электронным микроскопом миофибриллы представляют агрегаты из толстых, или миозиновых, и тонких, или актиновых, филаментов. Между толстыми филаментами располагаются тонкие филаменты ( диаметр 7-8 нм).
Толстые филаменты, или миозиновые нити, (диаметр 14 нм, длина 1500 нм, расстояние между ними 20-30 нм) состоят из молекул белка миозина, являющимся важнейшим сократительным белком мышцы, по 300-400 молекул миозина в каждой нити. Молекула миозина – это гексамер, состоящий из двух тяжелых и четырех легких цепей. Тяжелые цепи представляют собой две спирально закрученные полипептидные нити. Они несут на своих концах шаровидные головки. Между головкой и тяжелой цепью находится шарнирный участок, с помощью которого головка может изменять свою конфигурацию. В области головок – легкие цепи (по две на каждой). Молекулы миозина уложены в толстой нити таким образом, что их головки обращены наружу, выступая над поверхностью толстой нити, а тяжелые цепи образуют стержень толстой нити.
Миозин обладает АТФ-азной активностью: высвобождающаяся энергия используется для мышечного сокращения.
Тонкие филаменты, или актиновые нити, (диаметр 7-8 нм), образованы тремя белками: актином, тропонином и тропомиозином. Основным по массе белком является актин, который образует спираль. Молекулы тропомиозина располагаются в желобке этой спирали, молекулы тропонина располагаются вдоль спирали.
Толстые нити занимают центральную часть саркомера – А-диск, тонкие занимают I-диски и частично входят между толстыми миофиламентами. Н-зона состоит только из толстых нитей.
В покое взаимодействие тонких и толстых нитей (миофиламентов) невозможно, т.к. миозин-связывающие участки актина заблокированы тропонином и тропомиозином. При высокой концентрации ионов кальция конформационные изменения тропомиозина приводят к разблокированию миозин-связывающих участков молекул актина.
Двигательная иннервация мышечного волокна. Каждое мышечное волокно имеет собственный аппарат иннервации (моторная бляшка) и окружено сетью гемокапилляров, располагающихся в прилежащей РВСТ. Этот комплекс называется мион. Группа мышечных волокон, которые иннервируются одним мотонейроном, называется нервно-мышечной единицей. Мышечные волокна в этом случае могут располагаться не рядом (одно нервное окончание может контролировать от одного до десятков мышечных волокон).
Регенерация гладкой мышечной ткани. Гладкие миоциты характеризуются внутриклеточной регенерацией. При повышении функциональной нагрузки происходит гипертрофия миоцитов и в некоторых органах гиперплазия (клеточная регенерация). Так, при беременности гладко-мышечные клетки матки могут увеличиваться в 300 раз.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом