
Структурно-функциональная характеристика сердечной мышечной ткани. Источники развития и регенерации.
Собственно сердечная, мышечная ткань по своим свойствам занимает промежуточное положение между гладкими мышцами внутренних органов и поперечнополосатыми (скелетными). Она сокращается быстрее гладких, но медленнее поперечнополосатых мышц, работает ритмично и мало утомляется.
В связи с этим в ее строении имеется ряд своеобразных черт .Состоит эта ткань из отдельных мышечных клеток (миоцитов), почти прямоугольной формы, расположенных столбиком друг за другом. В целом получается структура, напоминающая поперечнополосатое волокно, разделенное на отрезки поперечными перегородками — вставочные диски,. Рядом лежащие волокна соединены анастомозами, что позволяет им сокращаться одновременно.
Структурно-функциональной единицей является клетка —кардиомиоцит. По строению и функциям кардиомиоциты подразделяются на две основные группы:
· типичные или сократительные кардиомиоциты, образующие своей совокупностью миокард;
· атипичные кардиомиоциты составляющие проводящую систему сердца и подразделяющиеся в свою очередь на три разновидности. Синусныекардиомиоциты передают управляющие сигналыпереходнымкардиомиоцитам, а последние-проводящим
Есть также .секреторные кардиомиоциты вырабатываютнатрийуретический фактор
Гистогенез Источники развития поперечно-полосатой мышечной ткани-симметричные участки висцерального листка спланхнотома в шейной части зародыша—миоэпикардиальные пластинки.Из них дифференцируются также клетки мезотелия эпикарда. В ходе гистогенеза возникает 5 видов кардиомиоцитов- рабочие, синусные переходные, проводящие, а также секреторные.
Возможности регенерации сердечной мышечной ткани: Стволовых клеток в сердечной мышце нет, поэтому погибающие кардиомиоциты не восстанавливаются.
Гладкая мышечная ткань. Структурная организация разновидностей гладких мышечных тканей. Структурные основы сокращения гладких мышечных клеток.. Иннервация.
Гладкая мышечная ткань образует достаточно большое количество важных внутренних органов в животном теле. Так, ей образованы: кишечник; половые органы; кровеносные сосуды всех типов; железы; органы выделительной системы; дыхательные пути; части зрительного анализатора; органы пищеварительной системы. –
Различают три группы гладких мышечных тканей- мезенхимные, эпидермальные и нейральные.
Структурная основа -Гладкий миоцит- веретеновидная клетка. Ядро палочковидное, находится в ее центральной части. Когда миоцит сокращается, его ядро изгибается и даже закручивается.
. Соседние миоцитов через отверстия в базальной мембране образуют друг с другом щелевидные сообщения (нексус), которые обеспечивают функциональные взаимодействия клеток.
Сократительных аппарат гладких миоцитов (миофибрилл) состоит из тонких миофиламентов, образованных актином, и толстым, сформированным миозином. Миоциты ограничены базальной мембраной, а также коллагеновыми (ретикулярными) эластичными волокнами
Эфферентная (моторная) иннервация гладких миоцитов осуществляется постганглионарными волокнами автономной нервной системы
Сокращение ГМТ медленное — тоническое, зато ГМТ малоутомляема.
ГМТ в эмбриональном периоде развивается из мезенхимы
Нервные ткани. Источники развития. Синапсы. Классификация синапсов.Строение, механизмы передачи импульсов
Нервная ткань— это система взаимосвязанных нервных клеток и нейроглии, обеспечивающих специфические функции восприятия раздражений, возбуждения, выработки импульса и передачи его.
Она является основой строения органов нервной системы, обеспечивающих регуляцию всех тканей и органов, их интеграцию в организме и связь с окружающей средой.
Нервные клетки(нейроны, нейроциты) — основные структурные компоненты нервной ткани, выполняющие специфическую функцию.
Нейроглия(neuroglia) обеспечивает существование и функционирование нервных клеток, осуществляя опорную, трофическую, разграничительную, секреторную и защитную функции.
Развитие. Нервная ткань развивается из дорсальной эктодермы. У 18-дневного эмбриона человека эктодерма формируетнервную пластинку, латеральные края которой образуютнервные валики, а между валиками формируется нервныйжелобок. Передний конец нервной пластинки образует головной мозг. Латеральные края образуютнервную трубку.Полость нервной трубки сохраняется у взрослых в виде системы желудочков головного мозга и центрального канала спинного мозга.
Читайте также: Основное свойство эпителиальной ткани
Синапсы– это структуры, предназначенные для передачи импульса с одного нейрона на другой или на мышечные и железистые структуры.
В зависимости от способа передачи импульсасинапсы могут быть химическими или электрическими (электротоническими).
Химические синапсы передают импульс на другую клетку с помощью специальных биологически активных веществ — нейромедиаторов, находящихся в синаптических пузырьках.
Область синаптического контакта между двумя нейронами состоит из пресинаптической мембраны, синаптической щели и постсинаптической мембраны.
Электрические, или электротонические, синапсы в нервной системе млекопитающих встречаются относительно редко. В области таких синапсов цитоплазмы соседних нейронов связаны щелевидными соединениями (контактами), обеспечивающими прохождение ионов из одной клетки в другую,
Вопрос 36. Поперечнополосатая сердечная мышечная ткань. Структурно-функциональная характеристика кардиомиоцитов.
Сердечная мышечная ткань образована клетками — кардиомиоцитами, связанными друг с другом в области вставочных дисков и образующими трехмерную сеть ветвящихся и анастомозирующих функциональных волокон. В отличии от скелетной, она представлена не симпластом, а синцитием. Кардиомиоциты — цилиндрические или ветвящиеся клетки, более крупные в желудочках. Кардиомиоциты содержат одно или два (в основном 2) ядра и саркоплазму, покрыта сарколеммой, которая снаружи окружена базальной мембраной. Ядра кардиомиоцитов — светлые, с преобладанием эухроматина, хорошо заметными ядрышками — занимают в клетке центральное положение.
Вставочные диски осуществляют связь кардиомиоцитов друг с другом. Под световым микроскопом они имеют вид поперечных прямых или зигзагообразных полосок. Под электронным микроскопом определяется сложная организация вставочного диска, представляющего собой комплекс межклеточных соединений нескольких типов: многочисленные интердигитации, десмосомы, полоски слипания, связь за счёт гликокаликса. На сарколемме продольных участков вставочного диска имеются многочисленные щелевые соединения, обеспечивающие ионную связь кардиомиоцитов и передачу импульса сокращения.
Кардиомиоциты бывают 5 видов:
Рабочие (сократительные) – образуют основную часть миокарда и характеризуются мощно развитым сократительным аппаратом, занимающим большую часть их саркоплазмы.
Синусные (пейсмейкерные) – за счёт них происходит создание импульса. Они занимают центральную часть синусного узла. В их цитоплазме наблюдается незначительное количество миофибрилл.
Проводящие – проводят импульс, образуя цепочки клеток, соединенных своими концами.
Переходные – служат промежуточным звеном между синусными и проводящими и между проводящими и рабочими.
Секреторные – вырабатывают натрийуретический фактор (гормон), участвующий в процессах регуляции мочеобразования и в некоторых других процессах.
Вопрос 37. Общая характеристика гладкой мышечной ткани. Типы гладких миоцитов. Строение, локализация и функции.
Гладкие мышцы выстилают некоторые внутренние органы, такие как кишечник, матка, мочевой пузырь, а также сосуды. Кроме того, из них формируются сфинктеры и связки. Структурной единицей гладкой мышечной ткани являются гладкие миоциты (ГМК), сократительный аппарат которых представлен не миофибриллами, а протофибриллами, которые также построены из актина и миозина. Структурной единицей протофибриллы является также саркомер.
По происхождению ГМК делятся на мезенхимные, эпидермальные и нейральные:
Читайте также: Наращивание костной ткани мази
Мышечная ткань мезенхимного происхождения. Гладкий миоцит – веретеновидная клетка длиной 20-500 мкм, шириной 5-8 мкм. Ядро палочковидное, находится в ее центральной части. Когда миоцит сокращается, его ядро изгибается и даже закручивается. Органеллы общего значения, среди которых много митохондрий, сосредоточены около полюсов ядра. Концы протофиламентов скреплены между собой и с плазмолеммой специальными сшивающими белками. Эти участки хорошо видны на электронных микрофотографиях как плотные тельца. Гладкие миоциты располагаются без заметных межклеточных пространств. На концах клеток плазмолемма образует узкие трубчатые впячивания. Миоциты разделены базальной мембраной. На отдельных участках в ней образуются «окна», поэтому плазмолеммы соседних миоцитов сближаются. Здесь формируются нексусы, и между клетками возникают не только механические, но и метаболические связи. Поверх «чехликов» из базальной мембраны между миоцитами проходят эластические и ретикулярные волокна, объединяющие клетки в единый тканевой комплекс.
Мышечная ткань эпидермального происхождения. Миоэпителиальные клетки развиваются из эпидермального зачатка. Они встречаются в потовых, молочных, слюнных и слезных железах и имеют общих предшественников с их секреторными клетками. Миоэпителиальные клетки непосредственно прилежат к собственно эпителиальным и имеют общую с ними базальную мембрану. При регенерации те и другие клетки тоже восстанавливаются из общих малодифференцированных предшественников. Большинство миоэпителиальных клеток имеют звездчатую форму. Эти клетки нередко называют корзинчатыми: их отростки охватывают концевые отделы и мелкие протоки желез. В теле клетки располагаются ядро и органеллы общего значения, а в отростках – сократительный аппарат, организованный, как и в клетках мышечной ткани мезенхимного типа
Мышечная ткань нейрального происхождения. Миоциты этой ткани развиваются из клеток нейрального зачатка в составе внутренней стенки глазного бокала. Тела этих клеток располагаются в эпителии задней поверхности радужки. Каждая из них имеет отросток, который направляется в толщу радужки и ложится параллельно ее поверхности. В отростке находится сократительный аппарат, организованный так же, как и во всех гладких миоцитах. В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы — суживающую и расширяющую зрачок.
Поперечно полосатая сердечная мышечная ткань структурно функциональная характеристика кардиомиоцитов
Гистогенез сердечной мышечной ткани. Источники развития сердечной мышечной ткани находятся в прекардиальной мезодерме. В гистогенезе возникают парные складчатые утолщения висцерального листка спланхнотома — миоэпикардиальные пластинки, содержащие стволовые клетки сердечной мышечной ткани. Последние путем дивергентной дифференцировки дают начало следующим клеточным дифферонам: рабочим, ритмзадающим (пейсмекерным), проводящим и секреторным кардиомиоцитам.
Исходные клетки сердечной мышечной ткани — кардиомиобласты характеризуются рядом признаков: клетки уплощены, содержат крупное ядро, светлую цитоплазму, бедную рибосомами и митохондриями. В дальнейшем происходит развитие комплекса Гольджи, гранулярной эндоплазматической сети. В кардиомиобластах обнаруживаются фибриллярные структуры, но миофибрилл нет. Клетки обладают высоким пролиферативным потенциалом. После ряда митотических циклов кардиомиобласты дифференцируются в кардиомиоциты, в которых начинается саркомерогенез. В цитоплазме кардиомиоцитов увеличивается число полисом, канальцев гранулярной эндоплазматической сети, накапливаются гранулы гликогена, возрастает объем актомиозинового комплекса. Кардиомиоциты сокращаются, но не теряют способность к дальнейшей пролиферации и дифференцировке. Развитие сократительного аппарата в позднем эмбриональном и постнатальном периодах происходит путем надставки новых саркомеров и наслоения вновь синтезированных миофиламентов. Дифференцировка кардиомиоцитов сопровождается увеличением числа митохондрий, распределением их у полюсов ядер и между миофибриллами и протекает параллельно со специализацией контактирующих поверхностей клеток. Кардиомиоциты путем контактов «конец в конец», «конец в бок» формируют клеточные комплексы — сердечные мышечные волокна, и в целом ткань представляет собой сетевидную структуру.
Читайте также: Ткани в кривом роге
Строение сердечной мышечной ткани.
Структурно-функциональные единицы волокон — кардиомиоциты — это клетки, имеющие вытянутую прямоугольную форму. Длина рабочих кардиомиоцитов составляет 50-120 мкм, а ширина — 15-20 мкм. Одно-два ядра располагаются в центре клетки. Периферическую часть цитоплазмы кардиомиоцитов занимают поперечноисчерченные миофибриллы, аналогичные таковым в симпластах скелетномышечного волокна. Однако каналы саркоплазматической сети и Т-системы менее отчетливо выражены. Кардиомиоциты отличаются большим количеством митохондрий, расположенных тесными рядами между миофибриллами. Снаружи миоциты покрыты сарколеммой, в составе которой выделяются плазмолемма и базальная мембрана. Характерной особенностью ткани является наличие вставочных дисков на границе между контактирующими кардиомиоцитами. Вставочные диски пересекают волокно в виде волнистой или ступенчатой линии и включают межклеточные контакты от простых, по типу десмо-сом и до щелевых (нексусов).
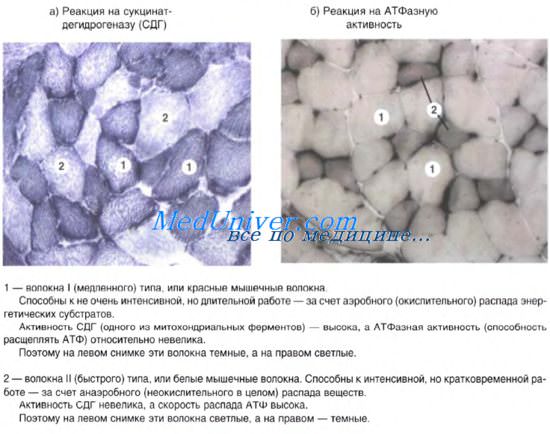
Часть кардиомиоцитов на ранних этапах кардиомиогенеза являются сократительно-секреторными. В дальнейшем в результате дивергентной дифференцировки возникают «темные» (сократительные) и «светлые» (проводящие) миоциты, в которых исчезают секреторные гранулы, тогда как в предсердных миоцитах они сохраняются. Так формируется дифферон эндокринных кардиомиоцитов. Эти клетки содержат центрально расположенное ядро с диспергированным хроматином,
1-2 ядрышками. В цитоплазме хорошо развиты гранулярная эндоплазматическая сеть, диктиосомы комплекса Гольджи, в тесной связи с элементами которого находятся многочисленные секреторные гранулы диаметром около 2 мкм, содержащие электронноплотный материал. В дальнейшем секреторные гранулы обнаруживаются под сарколеммой и выделяются в межклеточное пространство путем экзоцитоза. Выделенный пептидный гормон кардиодилатин циркулирует в крови в виде кардионатрина, который вызывает сокращение гладких миоцитов артериол, увеличение почечного кровотока, ускоряет клубочковую фильтрацию и выделение натрия из организма.
Кардиомиоциты проводящей системы гетероморфны. В них слабо развит мио-фибриллярный аппарат, расположение миофиламентов в составе миофибрилл рыхлое, Z-линии имеют неправильную конфигурацию, эндоплазматическая сеть слабо развита, находится на периферии миоцитов, число митохондрий незначительное. По мере расположения этих кардиомиоцитов в проксимо-дистальном направлении соответственно движению импульсов от синусно-предсердного узла, через предсердно-желудочковый узел, пучок Гиса, его ножки и клетки Пуркиня к рабочим миоцитам проводящие кардиомиоциты по своей ультраструктуре приближаются к рабочим кардиомиоцитам.
Регенерация сердечной мышечной ткани.
В гистогенезе сердечной мышечной ткани специализированный камбий не возникает. Поэтому регенерация ткани протекает на основе внутриклеточных гиперпластических процессов. Вместе с тем для кардиомиоцитов млекопитающих, приматов и человека характерен процесс полиплоидизации. Например, у обезьян ядра до 50% терминально дифференцированных кардиомиоцитов становятся тетра- и октоплоидными. Полиплоидные кардиомиоциты возникают за счет ацитокинетического митоза, что приводит к многоядерности.
В условиях патологии сердечно-сосудистой системы человека (ревматизм, врожденные пороки сердца, инфаркт миокарда и другие) важная роль в компенсации повреждений кардиомиоцитов принадлежит внутриклеточной регенерации, полиплоидизации как ядер, так и кардиомиоцитов.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом