
Мышечные ткани — это ткани, для которых способность к сокращению является главным свойством. Мышечные ткани составляют активную часть опорно-двигательного аппарата (пассивной частью являются кости, соединения костей).
Общими свойствами всех мышечных тканей является сократимость и возбудимость. К данной группе тканей относятся гладкая, поперечнополосатая скелетная и поперечнополосатая сердечная мышечные ткани. Клетки мышечной ткани имеют хорошо развитый цитоскелет, содержат много митохондрий.

Гладкая (висцеральная) мускулатура
Эта мышечная ткань встречается в стенках внутренних органах (бронхи, кишечник, желудок, мочевой пузырь), в стенках сосудов, протоках желез. Эволюционно является наиболее древним видом мускулатуры.
Состоит из веретенообразных миоцитов — коротких одноядерных клеток. Между клетками имеются межклеточные контакты — нексусы (лат. nexus — связь). Благодаря нексусам возбуждение, возникшее в одной клетке, волнообразно распространяется на все остальные клетки.

Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы внутренних органов (к примеру мочевого пузыря), сокращается медленно, практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает — сокращается и утомляется быстро.
Осуществляется сокращение с помощью клеточных органоидов — миофиламентов, которые расположены в клетке хаотично и не имеют такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их тоже изучим).
Особо заметим, что в гладкой мышечной ткани миофиламенты собираются в миофибриллы только во время сокращения. У таких временных миофибрилл не может быть регулярной организации, а значит ни у таких миофибрилл, ни у гладких миоцитов не может быть поперечной исчерченности.
Гладкая мышечная ткань сокращается непроизвольно (неподвластна воле человека). Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой. К примеру невозможно по желанию сузить или расширить бронхи, кровеносные сосуды, зрачок.

Гладкая мышечная ткань называется неисчерченной, так как не обладает поперечной исчерченностью, характерной для поперечнополосатых скелетной и сердечной мышечных тканей.
Скелетная (поперечнополосатая) мышечная ткань
Скелетная мышечная ткань образует диафрагму (дыхательную мышцу), мускулатуру туловища, конечностей, головы, голосовых связок.
В отличие от гладкой мускулатуры, скелетная образована не отдельными одноядерными клетками, а длинными многоядерными волокнами, имеющими до 100 и более ядер — миосимпластами. Миосимпласт (греч. sim — вместе + plast — образованный) представляет совокупность слившихся клеток, имеет длину от нескольких миллиметров до нескольких сантиметров (соответствует длине мышцы).
Внутри миосимпласта находится саркоплазма, снаружи миосимпласт покрыт сарколеммой. Сократительные элементы — миофибриллы (лат. fibra — волоконце) — длинные тяжеобразные органеллы в миосимпласте (около 1400).

Характерная черта данной ткани — поперечная исчерченность, выражающаяся в равномерном чередовании светлых и темных полос на мышечном волокне. Это происходит потому, что границы саркомеров в соседних миофибриллах совпадают, вследствие чего все волокно приобретает поперечную исчерченность. Теперь самое время изучить микроскопическую основу мышцы — саркомер.
Саркомер (от греч. sarco — мясо (мышца) + mere — маленький)
Саркомер — элементарная сократительная единица поперечнополосатых мышц, структурная единица миофибриллы. В состав саркомера (и миофибриллы в целом) входят миофиламенты (лат. filamentum — нить) двух типов, которые обеспечивают сократимость мышечной ткани.
Саркомер состоит из актиновых (тонких) и миозиновых (толстых) филаментов, которые образованы главным образом белками актином и миозином. Сокращение происходит за счет взаимного перемещения миофиламентов: они тянутся навстречу друг другу, саркомер укорачивается (и мышца в целом).

Источником энергии для сокращения служат молекулы АТФ. К тому же невозможно представить сокращение мышц без участия ионов кальция: именно они связываются с тропонином, что приводит к изменению конформации тропомиозина (тропонин и тропомиозин — регуляторные белки между нитями актина), за счет чего становится возможно соединение актина и миозина. При сокращении мышц выделяется тепло (сократительный термогенез).
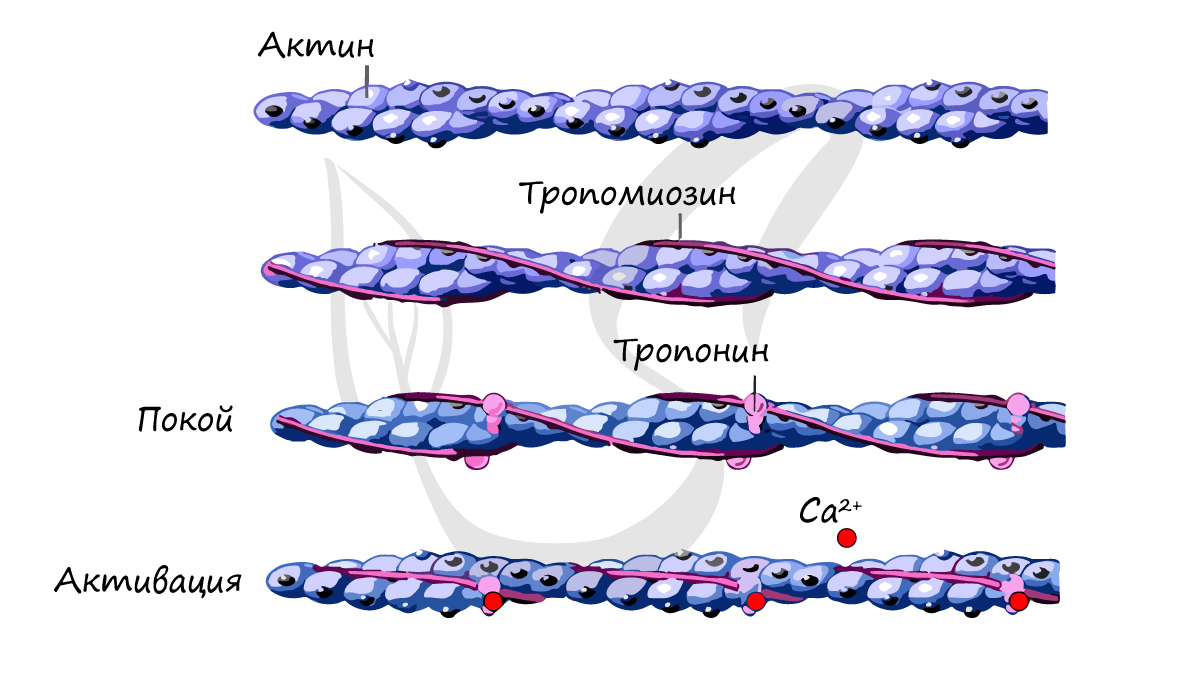
Замечу, что трупное окоченение (лат. rigor mortis) — посмертное затвердевание мышц — связано именно с ионами кальция, которые устремляются в область низкой концентрации (в саркоплазму миосимпласта), способствуя связыванию актина и миозина.
После смерти в мышце перестает синтезироваться АТФ, ее уровень быстро снижается. Как следствие этого перестает функционировать Ca-АТФаза — насос, выкачивающий ионы Ca из саркоплазмы в саркоплазматический ретикулум (мембранная органелла мышечных клеток (сходная с ЭПС), в которой запасаются ионы Ca).
В саркоплазме повышается концентрация ионов Ca — замыкаются мостики между актином и миозином, однако разомкнуться они уже не могут, в связи с чем наблюдается стойкая мышечная контрактура (лат. contractura — стягивание, сужение): конечности очень сложно разогнуть или согнуть.

Вернемся к скелетным мышцам. Имеется еще ряд важных моментов, о которых нужно знать.
В процесс возбуждения вовлекается изолированно один миосимпласт, соседние миосимпласты (волокна) не возбуждают друг друга, в отличие от гладких миоцитов, где возбуждение предается между соседними клетками через нексусы. Скелетные мышцы сокращаются быстро и быстро утомляются (у гладких мышц фазы сокращения и расслабления растянуты во времени, мало утомляются) .
Скелетные мышцы сокращаются произвольно: они подконтрольны нашему сознанию. К примеру, по желанию мы можем изменить скорость движения руки, темп бега, силу прыжка. Мышцы покрыты фасцией, крепятся к костям сухожилиями, и, сокращаясь, приводят в движение суставы.

Сердечная поперечнополосатая мышечная ткань
Сердечная мышечная ткань образует мышечную оболочку сердца — миокард (от др.-греч. μῦς «мышца» + καρδία — «сердце»). Миокард — средний слой сердца, составляющий основную часть его массы. При работе сердечная мышечная ткань не утомляется.

Сердечная мышечная ткань состоит из кардиомиоцитов — одиночных клеток, имеющих поперечную исчерченность. Соединяясь друг с другом, кардиомиоциты образуют функциональные волокна.
Читайте также: Особенности строения покровной ткани растений с функциями
Этот тип мышечной ткани удивительным образом сочетает свойства двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство — автоматизм.
Автоматизм — способность сердечной мышечной ткани возбуждаться и сокращаться самопроизвольно, без влияний извне. Это легко можно подтвердить, наблюдая сокращения изолированного сердца лягушки в физиологическом растворе: сокращения сердца в нем будут продолжаться несколько десятков минут после отделения сердца от организма.

Места контактов соседних кардиомиоцитов — вставочные диски (в их составе находятся нексусы), благодаря которым возбуждение одной клетки передается на соседние, таким образом волнообразно охватываются возбуждением и сокращаются новые участки миокарда.
Большое число контактов между кардиомиоцитами обеспечивает высокую эффективность и надежность проведения возбуждения по миокарду. Сокращается эта ткань непроизвольно, не утомляется.
На рисунке или микропрепарате узнать данную ткань можно по центральному положению ядер в клетках, поперечной исчерченности, наличию вставочных дисков и анастомозов (греч. anastomosis — отверстие) — мест соединений боковых поверхностей функциональных волокон (кардиомиоцитов).

В норме возбуждение проводится по проводящей системе сердца от предсердий к желудочкам (однонаправленно). Участок сердечной мышцы, в котором генерируются импульсы, определяющие частоту сердечных сокращений — водитель сердечного ритма.
Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных (англ. pacemaker — задающий ритм) клеток, которые также называют водителями ритма. Они спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.
Ответ мышц на физическую нагрузку
Физические нагрузки приводят к гипертрофии мышц (от др.-греч. ὑπερ- чрез, слишком + τροφή — еда, пища) — в них увеличивается количество мышечных волокон, объем мышечной массы нарастает.

В условиях гиподинамии (от греч. ὑπό — под и δύνᾰμις — сила), то есть пониженной активности, мышцы уменьшаются вплоть до полной атрофии (греч. а – «не» + trophe – питание). В худшем случае волокна мышечной ткани перерождаются в соединительную ткань, после чего пациент становится обездвиженным.

Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление. Гипертрофия сердца — состояние, требующее вмешательства врача и наблюдения за пациентом.
В большинстве случае гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы).

Происхождение мышц
Мышцы развиваются из среднего зародышевого листка — мезодермы.

© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Скелетная поперечно-полосатая мышечная ткань
Гистогенез. На определенном этапе развития миотомов сомитов распадаются на отдельные клетки — миобласты, которые состоят из одноядерных клеток цилиндрической или веретенообразной формы, с базофильной цитоплазмой. Часть миобластов остаются на месте, а часть выселяются в мезенхиму. При электронной микроскопии в цитоплазме миобластов обнаруживается эндоплазматическая сеть, состоящая из небольшого числа пузырьков с гладкой поверхностью. Митохондрии имеют овальную форму. Имеются многочисленные рибосомы. Миобласты дифференцируются в двух направлениях. Клетки одной линии сливаются , в результате чего образуются удлинененные симпластические структуры – миосимпласты, которые имеют неправильную форму, в центре концентрируются многочисленные ядра. Механизм образования миосимпластов является предметом продолжительной дискуссии. Большинство исследователей считают, что образование миосимпластов происходит в результате слияния нескольких миобластов. Однако последующее деление этих структур не завершается цитотомией, что приводит к образованию многоядерной структуры – миосимпласта. На этой стадии уже начинается образование миофибрилл на основе синтеза специфических сократительных белков (актина, миозина, тропомиозина). Первым признаком фибриллогенеза является образование в цитоплазме продольно ориентированных нитей, состоящих из мелких гранул. Собираясь в пучки эти нити формируют миофибриллы, различимые при световой микроскопии. Первые миофибриллы появляются в периферических участках миосимпластов уже у 5 –6 –недельных зародышей человека. Ведущая роль в синтезе сократительных белков принадлежит свободным рибосомам. Первоначально образовавшиеся миофибриллы тонкие и короткие. Рост их в толщину происходит за счет наслоения новых протофибрилл с периферии. Сначала протофибриллы лежат беспорядочно. В дальнейшем в результате роста миосимпластов в длину, увеличения в них числа миофибрилл и активного деления ядер образуются мышечные трубочки или миотубы. Они представляют собой длинные цилиндрической формы симпластические структуры с многочисленными ядрами, расположенными в центре в виде ядерно цепочки. При этом, миофибриллы располагаются по периферии. Развитие миотуб характеризуется постепенным увеличением числа миофибрилл. Часть миофибрилл возникает заново, но часть формируется при продольном расщеплении существующих после предварительного их утолщения. В результате увеличения числа миофибрилл они заполняют почти всю цитоплазму миотуб. При этом, ядра постепенно из центра перемещаются на периферию. Клеточные центры и микротрубочки полностью исчезают. Гранулярная сеть в значительной степени редуцируется. В результате происходит формирование мышечных волокон или миосимпластов. В период формирования дефинитивной структуры мышечных волокон их диаметр увеличивается. Это связано с гипертрофией саркоплазмы и значительным ростом числа миофибрилл. Одновременно волокна растут в длину. Процесс роста волокон более выражен в местах их перехода в сухожилие.
Клетки другой линии остаются самостоятельными и дифференцируются в миосателлитоциты (миосателлиты). Эти клетки лежат на поверхности (в углублениях) миосимпластов (мышечных волокон) и являются малодифференцированными клетками, являющимися источником регенерации мышечной ткани. Они прилежат к поверхности миосимпласта, так что их плазмолеммы соприкасаются. Миосимпласты и миосателлиты покрыты с поверхности общей базальной мембраной. Миосателлиты веретенообразной формы, одноядерные, их ядра овальной формы и более мелкие, чем в симпластах Эти клетки обладают всеми органеллами общего назначения, но развиты они слабо.Число клеток сателлитов составляет около 10% от числа ядер в мышечном волокне.
Читайте также: Pu полиуретановое покрытие для ткани
Структурно-функциональной единицей поперечно- полосатой мышечной ткани является мышечное волокно, которое представляет собой симпластическую структуру. Мышечное волокно имеет вид длинного цилиндрического образования, поперечник которого колеблется от 9 до 150 мкм, а длина достигает 12 и более см. Концы мышечных волокон закруглены или скошены. Иногда встречаются мышечные волокна с разветвленными концами.
Поверхность мышечного волокна покрыта тонкой оболочкой – сарколеммой, которая состоит из двух листков. Внутренний листок представляет собой плазматическую мембрану. Наружный листок является беструктурной мембраной, аналогичной базальной мембране, содержащей гликозаминогликаны. Между двумя листками плазмолеммы лежат многочисленные миосателлиты. К наружной поверхности сарколеммы прикрепляются коллагеновые фибриллы, которые осуществляют связь с с окружающей соединительной тканью. От сарколеммы внутрь волокна образуются впячивания. Они идут перпендикулярно к поверхности волокна. Это Т-каналы, которые обладают регидностью (жесткостью, негибкостью). По Т-каналам в цитоплазму волокна поступают вода и другие вещества.
В цитоплазме мышечного волокна (саркоплазме) под сарколеммой лежат многочисленные ядра. В зрелом мышечном волокне ядра имеют вытянутую форму. Их количество в одном симпласте может достигать нескольких десятков тысяч. В цитоплазме мышечного волокна содержится эндоплазматическая сеть, которая состоит из краевых цистерн, оплетающих Т-каналы, и центральной части, образованной продольными ветвящимися и анастомозирующимися друг с другом каналами с относительно узким просветом, получившим название L-каналы. Мембраны эндоплазматической сети не содержат рибосом. Митохондрии (саркосомы) в мышечных волокнах очень многочисленны. Их больше в быстро сокращяющихся волокнах. В периферической саркоплазме вблизи полюсов ядер располагается аппарат Гольджи и небольшие фрагменты гранулярной эндоплазматической сети.
Специальными органоидами в мышечном волокне являются миофибриллы. Они представляют собой тонкие нити, идущие по длине волокна. Число их в разных мышечных волокнах неодинаково. Каждая миофибрилла окружена продольно расположенными и анастомозирующими между собой петлями агранулярной эндоплазматической сети (саркоплазматической сети), в которой депонируются ионы кальция .Миофибриллы состоят в свою очередь из более тонких нитей – протофибрилл, состоящих из сократительных белков. При биохимическом исследовании установлено, что миофибриллы состоят из сократительных белков (80%), в том числе миозина (50%), актина (25%) и тропомиозина (5%). Различают два вида протофибрилл: миозиновые и актиновые. Миозиновые протофибриллы более толстые и состоят из белка миозина, а актиновые протофибриллы более тонкие и состоят из белка актина. Однотипные протофибриллы (и актиновые, и миозиновые) лежат упорядоченно, то есть параллельно друг другу. В результате упорядоченного расположения миозиновых протофибрилл формируются темные участки, обладающие двойным лучепреломлением, которые получили название анизотропных дисков (А- диски). Тонкие актиновые протофибриллы также располагаются упорядоченно, благодаря чему формируют светлые участки, обладающие одинарным лучепреломлением. Эти участки получили название изотропных дисков (И –диски). Чередование анизотропных и изотропных дисков обусловливает поперечную исчерченность мышечного волокна.При этом каждая миозиновая протофибрилла окружена 6 актиновыми протофибриллами. Посередине светлого диска располагается сеть из белковых фибриллярных молекул (прежде всего молекул белка альфа-актинина), с которой связаны концы актиновых филаментов. Эта сеть получила название Z-линии (телофрагмы, Т-линии). Филаменты актина объединены с Z-линией и нитями миозина фибриллярными нерастяжимыми молекулами небулина. Посередине темного анизотропного диска располагается фибриллярная сеть, построенная из миомезина. Она образует мезофрагму или М-линию. В узлах М-линии закреплены концы миозиновых филламентов. Другие их концы направляются в сторону Z-линии и располагаются между филламентами актина, но до самих Z-линий тоже не доходят. Вместе с тем эти концы фиксированы по отношению к Z- линии растяжимыми гигантскими белковыми молекулами титина. Участок миофибрилл от одной полоски Т до другой полоски называется саркомером, то есть соседние саркомеры имеют общую пограничную структуру – Z-линию. Саркомеры являются структурно-функциональными единицами миофибриллы.
Молекулы миозина имеют длинный хвост и на одном из его концов располагаются две головки. При повышении концентрации ионов кальция в области присоединения головок молекула изменяет свою конфигурацию. При этом, миозиновые головки связываются с актином (при участии тропомиозина и тропонина). После чего головка миозина наклоняется и тянет за собой актиновую молекулу в сторону М-линии. При этом, Z-линии сближаются и саркомер укорачивается.
Альфа-актиновые сети Z-линий соседних миофибрилл связаны друг с другом промежуточными филаментами. Они подходят к внутренней поверхности плазмолеммы и закрепляются в кортикальном слое цитоплазмы, так что саркомеры всех миофибрилл лежат на одном уровне. Это и создает поперечную исчерченность.
Когда клетка получает сигнал о начале сокращения, он перемещается по плазмолемме в виде потенциала действия и доходит до Т-трубочек. Поскольку эта мембрана сближена с мембранами саркоплазматического ретикулума, состояние последних меняется, кальций освобождается из цистерн сети и взаимодействует с актино-миозиновыми комплексами (они сокращаются). Когда потенциал действия исчезает, кальций снова аккумулируется и сокращение миофибрилл прекращается. Для сокращения нужна энергия, которая освобождается за счет расщепления АТФ ферментом АТФ-азой. Роль Атф-азы выполняет миозин. Источником АТФ служат главным образом митохондрии, поэтому они и располагаются непосредственно между миофибриллами. Большую роль в деятельности мышечных волокон играют включения миоглобина и гликогена. Гликоген служит источником энергии, необходимой не только для сокращения мышечной работы, но и поддержания теплового баланса всего организма. Так, установлено, что гликоген в саркоплазме расщепляется до глюкозы, а затем последняя расщепляется до молочной кислоты (гликолиз, то есть в отсутствии кислолрода), которая в присутствии кислорода расщепляется до углекислого газа и пировиноградной кислоты (дыхательный цикл Кребса). При этом освобождается АТФ. Часть энергии используется для превращения оставшейся молочной кислоты в гликоген. Миоглобин – это белок, по химическому строению очень близок к гемоглобину эритроцитов и тоже способен связывать кислород и отдавать его по мере надобности. Миоглобин связывает кислород, когда мышца расслаблена и через мелкие кровеносные сосуды свободно протекает кровь. Во время сокращения мышцы сосуды сдавливаются, а запасенный кислород освобождается и участвует в биохимических реакциях. Окраска миоглобина варьирует от красной до коричневой. Например, коричневая окраска мышцы тюленя обусловлена высоким содержанием миоглобина.
Читайте также: Ткани нетканые текстильные материалы товароведение
По содержанию миоглобина и на основе гистохимического анализа ферментативной активности и функциональных особенностей выделяют 3 типа (основных) мышечных волокон: белые (тип А), красные (тип В) и промежуточные (тип С).
Белые мышечные волокна более крупные, содержат мало миоглобина, миофибриллы лежат упорядоченно, содержат много гликогена и мало липидов, мало митохондрий, в них преобладают гликолитические процессы, низкая активность окислительно-восстановительных энзимов. Белые мышечные волокна быстро сокращаются и быстро расслабляются (утомляются),поэтому их называют тетоническими..
Красные мышечные волокна небольшого диаметра, содержат много миоглобина, миофибриллы лежат менее упорядоченно, содержат мало гликогена и много липидов, много митохондрий, в них преобладают окислительно-восстановительные процессы, но гликолитические процессы обладают низкой активностью. Эти волокна медленно сокращаются и медленно расслабляются, поэтому их называют тоническими.
Таким образом, по составу ферментов белые мышцы приспособлены к анаэробному гликолизу, а красные волокна – к окислительному обмену.
У человека и приматов эти типы мышечных волокон менее различимы, чем у других млекопитающих. У эмбриона первоначально все мышечные волокна являются красными, а затем появляются белые. Уже у плода человека 13 –17 недель указанные типы мышечных волокон вполне различимы. Свойства мышечных волокон меняются при изменении нагрузок – спортивных, профессиональных, а также в экстремальных условиях (невесомость). Эти изменения являются обратимыми. У диких птиц, способных к длительному полету, грудные мышцы содержат красные волокна, в то время как у домашних кур (нелетающих) грудные мышцы содержат белые волокна. В организме человека нет мышц, которые состоят из одного типа мышечных волокон, но есть мышцы, в которых преобладают те или другие.
Мышечные волокна, как правило, объединяются в пучки, образуя мышцы. В состав мышцы входит более 1миллиона мышечных волокон. При этом каждое мышечное волокно окружено тонкой прослойкой рыхлой неоформленной соединительной ткани и называется эндомизием. Коллагеновые волокна наружного листка сарколеммы (базальной мембраны) вплетаются в эндомизий, что способствует объединению усилий при сокращении миосимпластов (мышечных волокон). Группы мышечных волокон разграничиваются более выраженными прослойками рыхлой соединительной ткани – перимизий. В составе эндомизия и перимизия находится большое количество кровеносных и лимфатических сосудов, и нервов, за счет которых они выполняют трофическую функцию. Вокруг сосудов лежат многочисленные тучные клетки, принимающих участие в регуляции проницаемости сосудистой стенки.Поверхность мышцы покрыта наружным перимизием (эпимизием), который обеспечивает движение относительно соседних мышц с минимальной силой трения и кроме того выполняет опорную функцию.
Поперечно-полосатая мышечная ткань человека при повреждении способна к регенерации. Однако для полного восстановления необходим ряд условий, которые могут быть соблюдены только в эксперименте. В обычных условиях регенерация скелетной мышечной ткани, как правило, подавляется за счет разрастания соединительной ткани эндомизия и перимизия. Последняя быстро заполняет область дефекта и в конечном итоге образует в области краев поврежденных участков соединительнотканный рубец.
Восстановление скелетной мышечной ткани осуществляется за счет двух механизмов: компенсаторной гипертрофии самого симпласта и пролиферации миосателлитов. Ядра мышечных волокон делиться не могут, так как у них отсутствуют клеточные центры, поэтому камбиальными элементами служат миосателлиты. Пока организм растет, миосателлиты делятся, а дочерние клетки встраиваются в концы мышечных волокон. По окончании роста размножение миосателлитов затухает. При повреждении мышечного волокна на некотором расстоянии от места травмы оно разрушается и его фрагменты фагоцитируются макрофагами. В саркоплазме активируются аппарат Гольджи и гранулярная эндоплазматическая сеть, которые синтезируют необходимые вещества для восстановления саркоплазмы и миофибрилл. При этом, на поврежденном конце мышечного волокна формируется утолщение – мышечная почка. Миосателлиты под влиянием продуктов тканевого распада интенсивно пролиферируют и мигрируют к мышечной почке и встраиваются в нее. Часть миосателлитов сливаются (так же, как миобласты при гистогенезе) и образуют миотубы, которые формируют новые волокна, сливающихся с концами сохранившихся мышечных волокон и постепенно замещают дефект между ними. Процесс пролиферации миосателлитов регулируется рядом факторов, в том числе инсулиноподобным фактором роста, тромбоцитарным фактором роста, фактором роста фибробластов и др. Кроме того, одним из условий полноценной регенерации скелетных мышечных волокон является сохранение их базальной мембраны (наружного листка сарколеммы). Предполагают, что сохраненная базальная мембрана служит своеобразным барьером, предотвращающим проникновение клеток фибробластического ряда, но пропускающим макрофаги, поглощающие некротизированную ткань.
В настоящее время разрабатываются методы стимуляции восстановления структуры поврежденных мышц путем введения в них миосателлитов, так как разработаны методы выделения этих клеток из скелетных мышц. Кроме того, в настоящее время предпринята попытка использования этих клеток для замещения погибщей сердечной мышечной ткани. При этом, введенные миосателлиты в поврежденную середечнцю мышцу формируют волокна скелетной мышечной ткани, которые устанавливают связь с кардиомиоцитами.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом