
Существует ряд экспериментальных данных, свидетельствующих о том, что транскрипция осуществляется в так называемых транскрипционных фабриках: огромных, по некоторым оценкам, до 10 МДа комплексах, которые содержат около 8 РНК-полимераз II и компоненты последующего процессинга и сплайсинга, а также пруф-ридинга новосинтезированного транскрипта. В ядре клетки происходит постоянный обмен между пулами растворимой и задействованной РНК-полимеразы. Активная РНК-полимераза задействована в таком комплексе, который в свою очередь является структурной организовывающей компактизацию хроматина единицей. Последние данные. свидетельствуют о том, что транскрипционные фабрики существуют и в отсутствие транскрипции, они фиксированы в клетке (пока не ясно, взаимодействуют ли они с матриксом клетки или нет) и представляют собой независимый ядерный субкомпартмент. Попытки выделить белковый функциональный комплекс транскрипционной фабрики пока не привели к успеху ввиду его огромных размеров и низкой растворимости.
Транскрипция у эукариот
РНК-полимеразы эукариот
У эукариот имеется 3 типа РНК-полимераз (не считая митохондриальной и хлоропластной):
РНК полимеразаI — синтезирует в ядрышках рибосомные RNA (18S и 28S рРНК, кроме 5S);
РНК-полимеразаII — синтезирует mRNA и некоторых sRNA;
РНК-полимеразаIII — синтезирует tRNA, sRNA, 5S rRNA.
RNA-полимеразы эукариот отличаются: количеством субъединиц – 2 большие (120-220кДа) и до 8 малых (10-100кДа), потребностью в ионах Mg и Mn, чувствительностью к – амонитину — токсину бледной поганки — пептиду включающему D-аминокислоты: polI — устойчива, polII — ингибируется при концентрации 10-8М, polIII — при 10-6М амонитина. РНК-полимеразы I,II,III кодируются в ядре. Большие субъединицы гомологичны β и β`-субъединицам эубактерий.
РНК-полимераза I
РНК-полимераза II
PolII Человека содержит более 10 субъединиц, слабо ассоциирующих друг с другом. Некоторые из них принадлежат к основным факторам транскрипции (GTF).
Белки holo-фермента PolII дрожжей [Патрушев, 2000].
Pol II — РНК-Полимеразная активность, взаимодействует с множеством общих и тканеспецифических факторов транскрипции, участвует в выборе точки инициации транскрипции.
TFIIB — Связывает Pol II и TBP на промоторе, участвует в выборе точки инициации транскрипции
TFIIF — Взаимодействует с Pol II, стимулирует элонгацию транскрипции Pol II, компонент субкомплекса SRB/медиатор
TFIIH — Активность ДНК-зависимой ATPазы, ДНК-геликазная активность, обладает активностью CTD-киназы
SRB2, SRB5 — Участвуют в образовании инициационного комплекса, стимулируют базальный и индуцированный синтез РНК,
взаимодействуют с TBP, компоненты субкомплекса SRB/медиатор
GAL11/SPT13 — Участвуют в образовании инициационного комплекса, стимулируют базальный и индуцированный синтез РНК,
компоненты субкомплекса SRB/медиатор, предположительно взаимодействуют с активаторами транскрипции
SUG1 — Компонент субкомплекса SRB/медиатор, предположительно взаимодействует с активаторами транскрипции
SRB4, SRB6, SRB7, SRB8, SRB9, SRB10, SRB11 — Компоненты субкомплекса SRB/медиатор, предположительно
взаимодействуют с CTD-доменом Pol II
РНК-полимераза III
Факторы транскрипции
Инициация
Инициация транскрипции происходит на кэп-сайте кодирующем первый нуклеодид первого экзона мРНК.
ТАТА-бокс локализуется в 25-30 пн выше кэп-сайта связывая РНК-полимеразу перед кэп-сайтом. Промотор — примерно 200 пн выше кэп-сайта. Энхансеры обычно имеют длину 100-200 пн.
Элонгация
Терминация
Терминация на сайте полиаденилирования.
Вновь синтезированная РНК генов связывается с ядерными белками — информомерами, подвергается различным посттранскрипционным модификациям и транспортируется из ядра (см. обзор Процессинг) для последующей трансляции (см. обзор Трансляция).
Транскрипция у прокариот
РНК-полимераза E.coli
РНК-полимераза E.coli осуществляет транскрипцию всех бактериальных генов и состоит из нескольких субъединиц: α-35кДа, β‘-165кДа, β-155кДа, σ-чаще 70кДа (σ70). РНК-полимераза состава ααββ’σ70 называется holo-фермент (Еσ70), состава ααββ’- core-фермент (E).
σ — сменный фактор специфичности, который диссоциирует после инициации транскрипции. Элонгация и терминация осуществляется core-ферментом. У Е.coli
Читайте также: Перчатки для защиты рук из ткани
10 видов σ-субъединиц. Транскрипция генов теплового шока, оперонов gln или nif осуществляется σ54 в составе holo-фермента Eσ54 (54 кДа).
Все субъединицы заряжены отрицательно: σ>α>β>β’ – расположены по убыванию заряда. В каждой субъединице имеется кластер (+)-заряженных участков, которыми они связываются с ДНК. Наибольшее число кластеров у – β’, который участвует в связывании фермента с ДНК, β-субъединица содержит активные центры — инициации и элонгации, α-субъединицы обеспечивают правильное взаимодействие фермента с промоторами. Рифампицин – блокирует инициацию, стрептолидигин – блокирует элонгацию, что говорит о разнесении активных центров в РНК-полимеразе.
Узнавание и связывание RNA-pol с промотором осуществляется holo-ферментом
Одновременно в клетке присутствует около 7000 молекул РНК-полимеразы. Только holo-фермент обладает высоким сродством к специфической последовательности нуклеотидов — промотору, сродство к остальным случайным последовательностям ДНК у него снижено в 10000 раз. У core-фермента одинаковое сродство к любой последовательности нуклеотидов.
Сам по себе сигма — фактор обладает наименьшим сродством к ДНК по сравнению с другими субьединицами РНК-полимеразы, однако он придает holo-ферменту такую конформацию, которая обладает повышенным сродством к промотору.
Стадии узнавания и связывания, а также инициации осуществляются holo-ферментом. Элонгация и терминация осуществляются core-ферментом.
Две α субъединицы — каркас РНК-полимеразы. К ним крепятся остальные субъединицы.
β’ — субъединица отвечает за прочное связывание с ДНК за счет кластера положительно заряженных аминокислот.
В β — субъединице находятся два каталитических центра. Один отвечает за инициацию, а другой — за элонгацию. Один центр работает в holo-, а другой — в core- ферменте.
Инициация транскрипции
РНК-полимераза Ecoli узнает два 6н разделенных 25н
Элонгация транскрипции
Терминация транскрипции
Регуляция транскрипции
Схема негативной индукции Жакоба и Моно
Lac-оперон E. coli содержит 3 гена, отвечающие за образование белков, участвующих в переносе в клетку дисахарида лактозы и в ее расщеплении.
Z-β — галактозидаза (расщепляет лактозу на глюкозу и галактозу).
Y-β- галактозидпермеаза (переносит лактозу через мембрану клетки).
А — тиогалактозидтрансацетилаза (ацетилирует галактозу).
В отсутствие в клетке лактозы lac-оперон выключен. Активный белок — репрессор, кодируемый в моноцистронном опероне (LacI) , не имеющем оператора, связан с оператором lac-оперона. Поскольку оператор перекрывается с промотором, даже посадка РНК-полимеразы на промотор невозможна.
Как только некоторое количество лактозы попадает в клетку, две молекулы субстрата (лактозы) взаимодействуют с белком — репрессором, изменяют его конформацию — и он теряеет сродство к оператору.
Тут же начинается транскрипция lac-оперона и трансляция образующейся mРНК; три синтезируемых белка участвуют в утилизации лактозы.
Когда вся лактоза переработана, очередная порция репрессора, свободного от лактозы, выключает lac-оперон.
Схема позитивной индукции
Когда в клетку попадает арабиноза, она взаимодействует с белком — репрессором. Белок — репрессор меняет конформацию и превращается из репрессора в активатор, взаимодейсивующий с промотором и облегчающий посадку РНК-полимеразы на промотор.
Эта схема регуляции называется позитивной индукцией, поскольку контролирующий элемент — белок — активатор «включает» работу оперона.
Читайте также: Ткань для обивки диванов в красноярске
Схема позитивной репрессии
В опероне синтеза рибофлавина у Вacilus subtilis располагаются цистроны ферментов синтеза рибофлавина. Есть белок-активатор, обеспечивающий посадку РНК-полимеразы на промотор. В норме оперон открыт. Образуется N молекул рибофлавина.
N+1-ая молекула (лишняя) взаимодействует с активатором и он теряет способность активировать посадку РНК-полимеразы на промотор.
Позитивная репрессия, поскольку в регуляции участвует белок — активатор, а сама регуляция заключается в выключении транскрипции.
Схема негативной репрессии
В опероне синтеза триптофана у E. сoli имеется 5 цистронов, которые кодируют ферменты последовательной цепи реакций синтеза триптофана. В норме оперон включен. Белок — репрессор неактивен (в форме апо-репрессора), он не способен садиться на оператор.
Клетке нужно N молекул триптофана. N+1-ая молекула взаимодействует с апо-репрессором. Он меняет конформацию, садится на оператор и синтез РНК прекращается.
Схема регуляции — негативная репрессия, потому что белок репрессор «выключает» оперон.
Позитивный контроль работы lac-оперона
Lac-оперон, подчиняющийся схеме негативной индукции, имеет и позитивный контроль. цАМФ образуется из АТФ ферментом аденилатциклазой. Фосфодиэстераза превращает цАМФ в АМФ. Глюкоза активирует второй и инактивирует первый фермент. Чем больше в клетке глюкозы, тем меньше цАМФ.
Если нет глюкозы, то цАМФ соединяется с белком катаболической репрессии (САР) и образуется комплекс САР•цАМФ, активирующий посадку РНК-полимеразы на промотор. В присутствии лактозы lac-оперон включается и работает. Если же в клетке есть еще и глюкоза (более экономичный источнок энергии), то нет цАМФ — и активатор не образуется, lac-оперон работает слабо, без дополнительной индукции.
Этапы транскрипции
1. Инициация – первый этап транскрипции, в ходе которого происходит связывание РНК-полимеразы с промотором и образование первой межнуклеотидной связи.
У бактерий холофермент РНК-полимераза непосредственно узнает определенные последовательности нуклеотидных пар в составе промотора: последовательность 5-ТАТААТ-3 (расположена на расстоянии 10 нуклеотидов от точки начала транскрипции и называется боксом Прибнова) и последовательность 5-ТТГАЦА-3 (удалена от точки начала транскрипции на 35 нуклеотидов). В некоторых оперонах, например в лактозном, необходимо предварительное взаимодействие с промотором дополнительного белка (САР изменяет структуру промотора, резко повышая его сродство к РНК-полимеразе).
РНК-полимеразы эукариот не способны самостоятельно связываться с промоторами транскрибируемых генов. В присоединении к транскриптонам РНК-полимераз принимают участие общие факторы транскрипции (TF). Они отличаются от σ-факторов прокариот тем, что могут связываться с ДНК независимо от РНК-полимеразы. Полимеразы I, II и III требуют присутствия разных факторов транскрипции, обозначаемых TF I, TF II и TF III соответственно. Промоторы эукариот устроены более сложно, чем прокариотические, и состоят из нескольких элементов. Из низ самым близким к точке начала транскрипции является ТАТА-домен, называемый также доменом Хогнесса. Затем следуют домены ЦААТ и ГЦ. Промоторы эукариот могут содержать различные комбинации этих элементов, но ни один из них не встречается во всех промоторах. Домен ЦААТ играет существенную роль в инициации транскрипции, ТАТА и ГЦ, по-видимому, выполняют вспомогательные функции.
Связавшись с промотором, РНК-полимераза вызывает локальную денатурацию ДНК, т. е. разделение цепей ДНК на протяжении примерно 15 нуклеотидных пар. Образуется транскрипционный «глазок». Первым в строящуюся цепь РНК включается пуриновый нуклеотид – АТФ или ГТФ, при этом все три его фосфатных остатка сохраняются. После образования первой фосфодиэфирной связи σ-фактор у бактерий теряет связь с ферментом, и оставшийся core-фермент начинает перемещаться по ДНК. РНК-полимераза эукариот после инициации транскрипции также теряет связь с транскрипционными факторами и перемещается по ДНК самостоятельно.
Читайте также: Препарация тканей что это
2. Элонгация – последовательное удлинение растущей цепи РНК. Перемещаясь вдоль двойной спирали ДНК, РНК-полимераза непрерывно раскручивает спираль впереди того участка, где происходит синтез РНК. На короткое время образуется так называемый открытый комплекс, внутри которого возникает РНК-ДНК-спираль длиной около 20 нуклеотидов
(рис. 30). Затем фермент (с помощью специального сайта) вновь закручивает
Рис. 30. Элонгация транскрипции
ДНК позади участка полимеризации. РНК-транскрипт выводится из комплекса через особый канал, свойственный РНК-полимеразе.
Скорость синтеза РНК у бактерий составляет около 30 нуклеотидов в секунду, однако она не постоянна и может несколько снижаться. Такие периоды называют паузами транскрипции.
Показано, что еще до образования гибрида РНК-ДНК РНК-полимераза переводит ДНК из В-формы в А-форму. В ней плоскости азотистых оснований не перпендикулярны оси спирали, а наклонены на 20 0 к перпендикуляру. Вероятно, это облегчает разъединение двух соседних азотистых оснований в цепи ДНК. Параметры РНК-ДНК-спирали также практически полностью идентичны характеристикам А-формы ДНК.
3. Терминация (окончание транскрипции) определяется особой нуклеотидной последовательностью ДНК, расположенной в зоне терминатора оперона.
В бактериальных оперонах выделяют два типа терминаторов:
— ρ (ро) — независимые терминаторы (I типа);
— ρ — зависимые терминаторы (II типа).
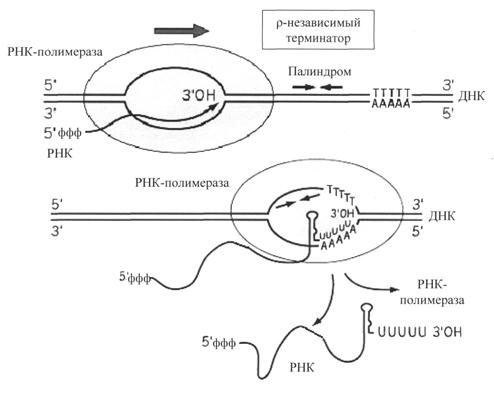
Рис. 31. ρ-независимая терминация транскрипции у бактерий
ρ-независимые терминаторы состоят из последовательностей, представляющих собой инвертированный повтор – палиндром (рис. 31), и располагаются за 16-20 нуклеотидных пар от точки терминации. Палиндромы (последовательности, которые читаются одинаково слева направо и справа налево) ρ-независимых терминаторов содержат большое количество Г-Ц-повторов. За этим участком на матричной цепи расположена олиго (А) — последовательность (4-8 адениловых нуклеотидов подряд). Транскрипция в области палиндрома приводит к тому, что в получившемся РНК-транскрипте быстро образуется устойчивый элемент вторичной структуры – «шпилька» – спирализованная область, содержащая комплементарные
Г-Ц-пары. «Шпилька» нарушает прочность связи ДНК-РНК в открытом комплексе. Кроме этого транскрипция олиго(А)-последовательности в матричной цепи ведет к образованию участка ДНК-РНК-гибрида, составленного из непрочных А-У пар, что также способствует разрушению контакта между ДНК и РНК.
ρ-зависимые терминаторы. Одним из факторов транскрипции прокариот является белок ρ. ρ-фактор – это имеющий четвертичную структуру белок, обладающий АТФ-азной активностью. Он способен связываться с 5-концом синтезируемой РНК длиной около 50 нуклеотидов. ρ-фактор движется по РНК с такой же скоростью, с которой РНК-полимераза движется по ДНК. Вследствие того что в терминаторе много Г-Ц-пар (с тремя водородными связями), РНК-полимераза в области терминатора замедляет ход, ρ-фактор ее догоняет, изменяет конформацию фермента, и синтез РНК прекращается (рис. 32).
На терминаторах обоих типов происходят три ключевых события:
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом