
В нервных волокнах сигналы передаются с помощью потенциалов действия, которые представляют собой быстрые изменения мембранного потенциала, быстро распространяющиеся вдоль мембраны нервного волокна. Каждый потенциал действия начинается со стремительного сдвига потенциала покоя от нормального отрицательного значения до положительной величины, затем он почти так же быстро возвращается к отрицательному потенциалу. При проведении нервного сигнала потенциал действия движется вдоль нервного волокна вплоть до его окончания.
На рисунке показаны изменения, возникающие на мембране во время потенциала действия, с переносом положительных зарядов внутрь волокна вначале и возвращением положительных зарядов наружу в конце. В нижней части рисунка графически представлены последовательные изменения мембранного потенциала в течение нескольких 1/10000 сек, иллюстрирующие взрывное начало потенциала действия и почти столь же быстрое восстановление.
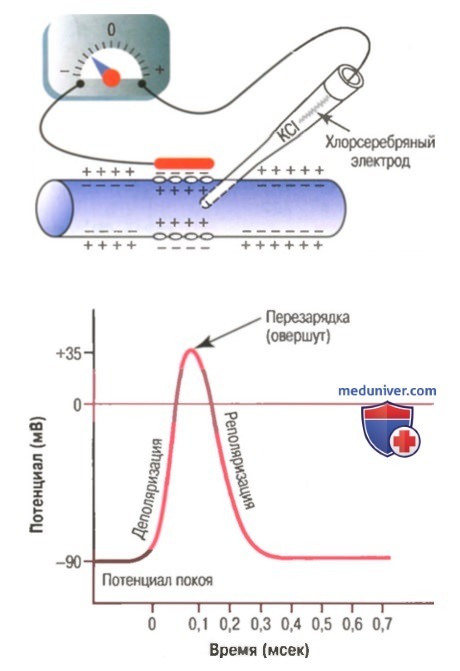
Типичный потенциал действия, зарегистрированный с помощью метода, представленного в верхней части рисунка.
Стадия покоя. Эта стадия представлена мембранным потенциалом покоя, который предшествует потенциалу действия. Мембрана во время этой стадии поляризована в связи с наличием отрицательного мембранного потенциала, равного -90 мВ.
Фаза деполяризации. В это время мембрана внезапно становится высокопроницаемой для ионов натрия, позволяя огромному числу положительно заряженных ионов натрия диффундировать внутрь аксона. Нормальное поляризованное состояние в -90 мВ немедленно нейтрализуется поступающими внутрь положительно заряженными ионами натрия, в результате потенциал стремительно нарастает в положительном направлении. Этот процесс называют деполяризацией, В крупных нервных волокнах значительный избыток входящих внутрь положительных ионов натрия обычно приводит к тому, что мембранный потенциал «проскакивает» за пределы нулевого уровня, становясь слегка положительным. В некоторых более мелких волокнах, как и в большинстве нейронов центральной нервной системы, потенциал достигает нулевого уровня, не «перескакивая» его.
Фаза реполяризации. В течение нескольких долей миллисекунды после резкого повышения проницаемости мембраны для ионов натрия, натриевые каналы начинают закрываться, а калиевые — открываться. В результате быстрая диффузия ионов калия наружу восстанавливает нормальный отрицательный мембранный потенциал покоя. Этот процесс называют реполя-ризацией мембраны.
Для более полного понимания факторов, являющихся причиной деполяризации и реполяризации, необходимо изучить особенности двух других типов транспортных каналов в мембране нервного волокна: электроуправляемых натриевых и калиевых каналов.
Электроупавляемые натриевые и калиевые каналы. Необходимым участником процессов деполяризации и реполяризации во время развития потенциала действия в мембране нервного волокна является электроуправляемый натриевый канал. Электроуправляемый калиевый канал также играет важную роль в увеличении скорости реполяризации мембраны. Оба типа электроуправляемых каналов существуют дополнительно к Na + /K + -насосу и каналам К + /Na + -утечки.
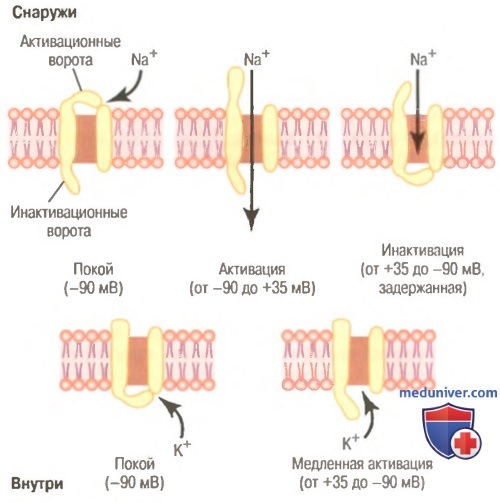
Типичные изменения состояния электроуправляемых натриевых (вверху) и калиевых (внизу) каналов при изменении мембранного потенциала от нормального отрицательного значения до положительных величин.
Показана последовательная активация и инактивация натриевых каналов и задержанная активация калиевых каналов.
Электроуправляемый натриевый канал. В верхней части рисунка показан электроуправляемый натриевый канал в трех различных состояниях. Этот канал имеет двое ворот: одни вблизи наружной части канала, которые называют активационными воротами, другие — у внутренней части канала, которые называют инактивационными воротами. В верхней левой части рисунка изображено состояние этих ворот в покое, когда мембранный потенциал покоя равен -90 мВ. В этих условиях активационные ворота закрыты и препятствуют поступлению ионов натрия внутрь волокна.
Активация натриевого канала. Когда мембранный потенциал покоя смещается в направлении менее отрицательных значений, поднимаясь от -90 мВ в сторону нуля, на определенном уровне (обычно между -70 и -50 мВ) происходит внезапное конформационное изменение актива-ционных ворот, в результате они переходят в полностью открытое состояние. Это состояние называют активированным состоянием канала, при котором ионы натрия могут свободно входить через него внутрь волокна; при этом натриевая проницаемость мембраны возрастает в диапазоне от 500 до 5000 раз.
Инактивация натриевого канала. В верхней правой части рисунке показано третье состояние натриевого канала. Увеличение потенциала, открывающее активационные ворота, закрывает инактивационные ворота. Однако инактивационные ворота закрываются в течение нескольких десятых долей миллисекунды после открытия активационных ворот. Это значит, что конформационное изменение, приводящее к закрытию инактивационных ворот, — процесс более медленный, чем конформационное изменение, открывающее активационные ворота. В результате через несколько десятых долей миллисекунды после открытия натриевого канала инактивационные ворота закрываются, и ионы натрия не могут более проникать внутрь волокна. С этого момента мембранный потенциал начинает возвращаться к уровню покоя, т.е. начинается процесс реполяризации.
Читайте также: Электропроводность живых тканей это
Существует другая важная характеристикая процесса инактивации натриевого канала: инактивационные ворота не открываются повторно до тех пор, пока мембранный потенциал не вернется к значению, равному или близкому к уровню исходного потенциала покоя. В связи с этим повторное открытие натриевых каналов обычно невозможно без предварительной реполяризации нервного волокна.
Видео физиология возбуждения тканей (потенциал покоя, потенциал действия) — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в оглавление раздела «Физиология человека.»
Общая физиология возбудимых тканей Лекция 1
Раздражимость – свойство всех живых клеток: способность отвечать на действие раздражителя (изменение метаболизма, движение, деление клеток и др.).
Возбудимость – способность клеток отвечать не действие раздражителя возбуждением (т.е.генерацией потенциала действия). К возбудимым тканям относятся нервная и мышечная.
Возбуждение – ответ возбудимой клетки на действие раздражителя (потенциал действия).
Раздражитель – изменение внешней или внутренней среды, которое действует на клетку и вызывает ответную реакцию. Раздражитель может быть: (1) химический, электрический, механический и др., (2) пороговый, сверхпороговый, подпороговый; (3) адекватный и неадекватный и т.д.
МЕМБРАННО-ИОННАЯ ТЕОРИЯ ВОЗБУЖДЕНИЯ.
Мембранные потенциалы создаются за счет движения ионов через клеточную мембрану.
Мембрана – двойной слой фосфолипидов – проницаема для жирорастворимых веществ (СО2, О2, спирт, эфир, стероидные гормоны и др.) и непроницаема для водорастворимых веществ, в том числе для ионов. Для движения ионов в мембране существуют специальные белковые структуры.
Ионные каналы – поры в мембране, стенки которых образованы белковыми молекулами. Через ионные каналы происходит диффузия ионов. Движущей силой для диффузии ионов является (1) концентрационный градиент и (2) электрический градиент. Диффузия происходит без затрат энергии АТФ и называется пассивным транспортом (движение ионов из области с большей концентрацией в область с меньшей концентрацией данных ионов). Ионные каналы бывают нерегулируемые (всегда открыты) и регулируемые (могут быть открыты или закрыты).
Ионные насосы – белковые молекулы-переносчики, которые обеспечивают активный транспорт ионов с затратами энергии АТФ (движение ионов из области с меньшей концентрацией в область с большей концентрацией ионов). Например, калий-натриевый насос (К-Na-АТФаза) имеется в мембране всех живых клеток и переносит ионы калия в клетку, а ионы натрия – из клетки. Поэтому в клетках всегда концентрация ионов калия выше, чем в тканевой жидкости, а концентрация ионов натрия к клетке всегда ниже, чем в тканевой жидкости. Функция К-Na-насоса – создавать и поддерживать градиенты концентраций ионов!
– разность потенциалов, которая существует между внутренней поверхностью клеточной мембраны (-) и наружной поверхностью клеточной мембраны (+) в покое. Эту разность потенциалов можно измерить с помощью микроэлектрода, который вводится в клетку (активный электрод), в то время как второй электрод большей площади остается во внеклеточной среде (пассивный электрод, нулевой, электрод сравнения). Потенциал покоя равен (-30 мв) – (-90 мв) в клетках разных тканей.
Механизм формирования ПП. Необходимы два условия: (1) разная концентрация ионов в клетке и в тканевой жидкости и (2) разная проницаемость мембраны для разных ионов.
В покое проницаемость клеточной мембраны для калия в десятки раз больше, чем для натрия. Поэтому происходит выход ионов калия из клетки (диффузия из большей концентрации в меньшую). Ионы калия, выходя из клетки, заряжают наружную поверхность мембраны положительно, а крупные органические анионы, оставаясь в клетке, заряжают внутреннюю поверхность мембраны отрицательно. Чем больше разность концентраций калия, тем больше разность потенциалов (закон Нернста).
Примечание: В покое проницаемость клеточной мембраны для натрия очень низкая. Тем не менее, это приводит к постоянной утечке ионов: небольшое количество ионов натрия проникает в клетку и вытесняет небольшое количество ионов калия из клетки. Утечке ионов противодействует постоянная работа К-Na-насосов: на 1 канал утечки приходится 100 насосных молекул (которые возвращают калий в клетку, а натрий вывадят из клетки). Если работу насосов прекратить, концентрации ионов выравниваются через несколько минут за счет утечки ионов. Мембранные потенциалы при этом равны нулю, функции клеток нарушаются.
– быстрое изменение мембранного потенциала под действием раздражителя.
Фазы потенциала действия: (а) деполяризация (медленная, затем быстрая), (б) инверсия потенциала, (в) реполяризация; затем могут быть следовые потенциалы (следовая деполяризация, следовая гиперполяризация.
Читайте также: Спиральная связка представлена тканью
Деполяризация – это уменьшение мембранного потенциала покоя (уменьшение величины отрицательного заряда на внутренней поверхности мембраны, напрмер, от -70 мв до 0).
Инверсия — изменение знака заряда на противоположный.
Реполяризация – восстановление исходного мембранного потенциала покоя.
Гиперполяризация – это увеличение мембранного потенциала по сравнению с уровнем ПП (увеличение отрицательного заряда на внутренней поверхности мембраны, например, от -70 мв до -100 мв)
Механизм возникновения ПД: (1) Под действием раздражителя увеличивается проницаемость клеточной мембраны для ионов натрия (открываются быстрые потенциал-чувствительные натриевые каналы). (2) Начинается диффузия ионов натрия в клетку за счет электрохимического градиента (пассивный транспорт). (3) Ток натрия в клетку вызывает деполяризацию клеточной мембраны, а затем и инверсию мембранного потенциала (до +30 мв). (4) К моменту инверсии потенциала натриевый ток прекращается (быстрые натриевые каналы инактивируются). (5) Увеличивается проницаемость клеточной мембраны для ионов калия (открываются медленные потенциал-чувствительные калиевые каналы). (6) Происходит диффузия ионов калия из клетки за счет электрохимического градиента (пассивный транспорт). (7) Ток калия из клетки вызывает реполяризацию и устанавливается потенциал покоя.
Продолжительность ПД – несколько миллисекунд; амплитуда ПД – 100-120 милливольт.
(Смотри схему ПД в учебнике, научись рисовать схему ПД самостоятельно!)
Восстановительный период: (а) восстанавливается исходный потенциал покоя; (б) восстанавливается исходное состояние натриевых и калиевых каналов; (в) восстанавливаются градиенты концентраций калия и натрия (за счет усиленной работы К-Na-насосов).
Быстрые потенциал-чувствительные натриевые каналы – имеют двое ворот: активационные и инактивационные. В покое активационные ворота закрыты, инактивационные открыты.Деполяризация вызывает быструю активацию натриевых каналов (открываются активационные ворота, инактивационные ворота тоже еще открыты). Канал открыт, натрий поступает в клетку. Затем происходит инактивация натриевых каналов, т.к. закрываются инактивационные ворота. За время реполяризации натриевые каналы должны вернуться в исходное состояние, характерное для ПП.
Медленные потенциал-чувствительные калиевые каналы – имеют только одни ворота. В покое эти ворота закрыты. Деполяризация вызывает медленную активацию калиевых каналов (открываются ворота). К концу реполяризации и сразу после нее ворота в калиевых каналах закрываются.
Потенциал действия
Потенциа́л де́йствия — волна возбуждения, перемещающаяся по мембране живой клетки в процессе передачи нервного сигнала. По сути своей представляет электрический разряд — быстрое кратковременное изменение потенциала на небольшом участке мембраны возбудимой клетки (нейрона, мышечного волокна или железистой клетки), в результате которого наружная поверхность этого участка становится отрицательно заряженной по отношению к соседним участкам мембраны, тогда как его внутренняя поверхность становится положительно заряженной по отношению к соседним участкам мембраны. Потенциал действия является физической основой нервного или мышечного импульса, играющего сигнальную (регуляторную) роль.

Рис. 1. Схема распределения зарядов по разные стороны мембраны возбудимой клетки в спокойном состоянии (A) и при возникновении потенциала действия (B) (см. объяснения в тексте).
Потенциалы действия могут различаться по своим параметрам в зависимости от типа клетки и даже на различных участках мембраны одной и той же клетки. Наиболее характерный пример различий: потенциал действия сердечной мышцы и потенциал действия большинства нейронов. Тем не менее, в основе любого потенциала действия лежат следующие явления:
Мембрана живой клетки поляризована — её внутренняя поверхность заряжена отрицательно по отношению к внешней благодаря тому, что в растворе возле её внешней поверхности находится бо́льшее количество положительно заряженных частиц (катионов), а возле внутренней поверхности — бо́льшее количество отрицательно заряженных частиц (анионов).
Мембрана обладает избирательной проницаемостью — её проницаемость для различных частиц (атомов или молекул) зависит от их размеров, электрического заряда и химических свойств.
Мембрана возбудимой клетки способна быстро менять свою проницаемостъ для определённого вида катионов, вызывая переход положительного заряда с внешней стороны на внутреннюю (Рис.1).
Первые два свойства характерны для всех живых клеток. Третье же является особенностью клеток возбудимых тканей и причиной, по которой их мембраны способны генерировать и проводить потенциалы действия.
Предспайк — процесс медленной деполяризации мембраны до критического уровня деполяризации (местное возбуждение, локальный ответ).
Пиковый потенциал, или спайк, состоящий из восходящей части (деполяризация мембраны) и нисходящей части (реполяризация мембраны).
Отрицательный следовой потенциал — от критического уровня деполяризации до исходного уровня поляризации мембраны (следовая деполяризация).
Читайте также: Обычный узел из ткани
Положительный следовой потенциал — увеличение мембранного потенциала и постепенное возвращение его к исходной величине (следовая гиперполяризация).
Общие положения

Рис. 2. A.Схематичное изображение идеализированного потенциала действия.B.Реальный потенциал действия пирамидного нейронагиппокампакрысы. Форма реального потенциала действия обычно отличается от идеализированной.
Поляризация мембраны живой клетки обусловлена отличием ионногосостава с её внутренней и наружной стороны. Когда клетка находится в спокойном (невозбуждённом) состоянии, ионы по разные стороны мембраны создают относительно стабильную разность потенциалов, называемуюпотенциалом покоя. Если ввести внутрь живой клеткиэлектроди измерить мембранный потенциал покоя, он будет иметь отрицательное значение (порядка −70 — −90 мВ). Это объясняется тем, что суммарный заряд на внутренней стороне мембраны существенно меньше, чем на внешней, хотя с обеих сторон содержатся икатионы, ианионы. Снаружи — на порядок большеионовнатрия,кальцияихлора, внутри — ионовкалияи отрицательно заряженныхбелковыхмолекул, аминокислот, органических кислот,фосфатов,сульфатов. Надо понимать, что речь идёт именно о заряде поверхности мембраны — в целом среда и внутри, и снаружи клетки заряжена нейтрально.
Потенциал мембраны может изменяться под действием различных стимулов. Искусственным стимулом может служить электрический ток, подаваемый на внешнюю или внутреннюю сторону мембраны через электрод. В естественных условиях стимулом часто служит химический сигнал от соседних клеток, поступающий черезсинапсили путёмдиффузнойпередачи через межклеточную среду. Смещение мембранного потенциала может происходить в отрицательную (гиперполяризация) или положительную (деполяризация) сторону.
В нервной ткани потенциал действия, как правило, возникает при деполяризации — если деполяризация мембраны нейрона достигает некоторого пороговогоуровня или превышает его, клетка возбуждается, и от её тела каксонамидендритамраспространяется волна электрического сигнала. (В реальных условиях на теле нейрона обычно возникают постсинаптические потенциалы, которые сильно отличаются от потенциала действия по своей природе — например, они не подчиняются принципу «всё или ничего». Эти потенциалы преобразуются в потенциал действия на особом участке мембраны —аксонном холмике, так что потенциал действия не распространяется на дендриты).
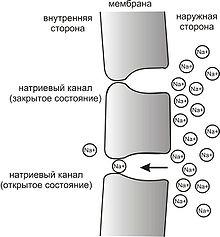

Рис. 3.Простейшая схема, демонстрирующая мембрану с двумя натриевыми каналами в открытом и закрытом состоянии, соответственно
Это обусловлено тем, что на мембране клетки находятся ионные каналы— белковые молекулы, образующие в мембране поры, через которые ионы могут проходить с внутренней стороны мембраны на наружную и наоборот. Большинство каналов ионоспецифичны — натриевый канал пропускает практически только ионы натрия и не пропускает другие (это явление называют селективностью). Мембрана клеток возбудимых тканей (нервной и мышечной) содержит большое количествопотенциал-зависимыхионных каналов, способных быстро реагировать на смещение мембранного потенциала. Деполяризация мембраны в первую очередь вызывает открытие потенциал-зависимых натриевых каналов. Когда одновременно открывается достаточно много натриевых каналов, положительно заряженные ионы натрия устремляются через них на внутреннюю сторону мембраны. Движущая сила в данном случае обеспечиваетсяградиентомконцентрации (с внешней стороны мембраны находится намного больше положительно заряженных ионов натрия, чем внутри клетки) и отрицательным зарядом внутренней стороны мембраны (см. Рис. 2). Поток ионов натрия вызывает ещё бо́льшее и очень быстрое изменение мембранного потенциала, которое и называютпотенциалом действия(в специальной литературе обозначается ПД).
Согласно закону «всё-или-ничего»мембрана клетки возбудимой ткани либо не отвечает на стимул совсем, либо отвечает с максимально возможной для неё на данный момент силой. То есть, если стимул слишком слаб и порог не достигнут, потенциал действия не возникает совсем; в то же время, пороговый стимул вызовет потенциал действия такой жеамплитуды, как и стимул, превышающий пороговый. Это отнюдь не означает, что амплитуда потенциала действия всегда одинакова — один и тот же участок мембраны, находясь в разных состояниях, может генерировать потенциалы действия разной амплитуды.
После возбуждения нейрон на некоторое время оказывается в состоянии абсолютной рефрактерности, когда никакие сигналы не могут его возбудить снова, затем входит в фазуотносительной рефрактерности, когда его могут возбудить исключительно сильные сигналы (при этом амплитуда ПД будет ниже, чем обычно). Рефрактерный период возникает из-за инактивации быстрого натриевого тока, то есть инактивации натриевых каналов (см. ниже).
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом