
Химизация сельскохозяйственного производства важнейший элемент повышения его продуктивности. Но давайте задумаемся, в погоне за высокой урожайностью не открывают ли технологи «ящик Пандоры», ведь последствия от применения малоизученных препаратов в долгосрочной перспективе могут быть самые неожиданные.
Одним из направлений химизации есть сбалансированное обеспечение растений всеми необходимыми макро и микроэлементами питания, которые активизируют процессы, необходимые для успешного развития растений и повышения их продуктивности. Однако последние должны вводится в живой организм в биологически активной форме, способной легко транспортироваться и усваиваться. Этого можно достичь при использовании биологически активных комплексонов. Наиболее распространенным примером таких комплексонов является ЭДТА (этилендиа- минтетрауксусная кислота), впервые полученная австрийским химиком Фердинандом Мюнцом (1888-1969 гг.) в Германии, который запатентовал ее в 1935 году. В современных технологиях производства ЭДТА сырьем для ее синтеза служат этилендиамин, формальдегид и синильная кислота, которые впоследствии подвергают щелочному гидролизу.
Основное свойство ЭДТА – это способность связывать ионы некоторых элементов,в число которых входят Са2+, Fe3+, Cu2+, Ni2+, Со2+,образовывая с ними особенно стабильные комплексы, называемые хелатами (от греческого chele – крабовая клешня).
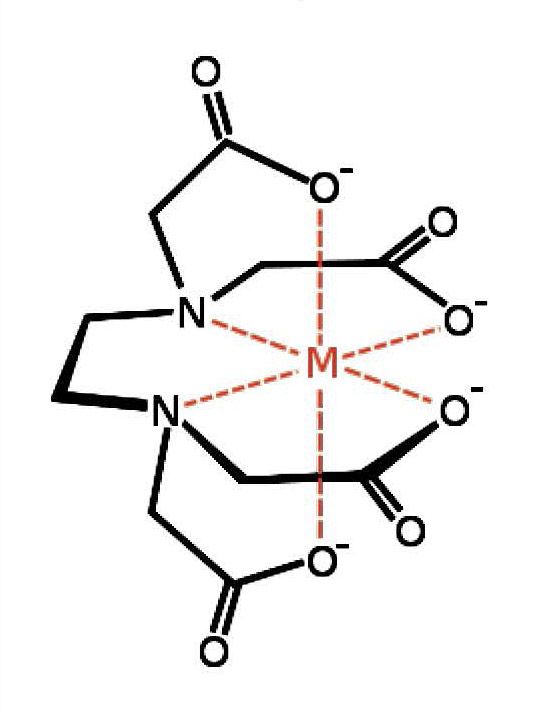
Эта способность, а также относительная дешевизна этого хелатанта обусловили использование его в различных областях деятельности человека, таких как аналитическая химия, медицина, промышленность, сельское хозяйство.
Для химических препаратов, применяемых в указанных отраслях, важнейшим свойством является их токсичность. Основной принцип медицины – «не навреди». Это в полной мере касается всех видов деятельности человека, которые так или иначе могут влиять на здоровье людей и на экологическую обстановку в целом. Все химические вещества, которые человек использует в сельском хозяйстве, пищевой промышленности и т. п., должны быть нетоксичными и не накапливаться в почве и живых организмах.
Отклонение от этих требований обязательно создаст через определенное время серьезные проблемы.
ЭДТА и коплексонаты на ее основе достаточно стабильны, что, с одной стороны, является преимуществом, а с другой – создает опасность накопления этого вещества в почве при систематическом его применении.
В чем же заключается угроза?
При попадании в растение комплексонат отдает атомы металлов (например, железа, марганца) растению, освобождая ЭДТА в чистом виде. Это вещество не усваивается растением и в конечном итоге попадает в почву, концентрируясь в основном в ее верхнем 10-сантиметровом слое.
Далее ЭДТА проявляет высокую устойчивость, увеличивая подвижность металлов в субстрате, продолжая выполнять возложенную на нее функцию и оказывая влияние на уровень накопления растениями металлов, даже на третий год после ее однократного внесения [Автухович И. Е.,Постников Д. А. Влияние ЭДТА на поведение металлов в субстрате и их накопление растениями //Естественные и технические науки. – 2014].
Так, через 19 месяцев после применения ЭДТА обнаруживается в корневой зоне, создавая, по сути, мину замедленного действия для будущей урожайности сельскохозяйственных культур, поскольку в последнее время в связи с бурным развитием промышленности наблюдается значительное возрастание уровня тяжелых металлов в окружающей среде.
Термин «тяжелые металлы» применяется к металлам либо с плотностью, превышающей 5 г/см3, либо с атомным номером больше 20. Хотя существует и другая точка зрения, согласно которой к тяжелым металлам относятся свыше 40 химических элементов с атомными массами, превышающими 50 ат. ед. Среди химических элементов тяжелые металлы наиболее токсичны и уступают по уровню своей опасности только пестицидам. При этом к токсичным относятся следующие химические элементы: Co, Ni, Cu, Zn, Sn, As,Se, Te, Rb, Ag, Cd, Au, Hg, Pb,Sb, Bi, P.
Тяжелые металлы, находящиеся в грунте в виде солей, слабо усваиваются растениями. Но образование комплексов ЭДТА с тяжелыми металлами приводит к более интенсивному поглощению их растениями, что бывает крайне нежелательно.
Тяжелые металлы токсичны для растений, их концентрация приводит к хлорозу, слабому росту растений, снижению урожайности и может даже сопровождаться снижением поглощения питательных веществ, нарушением в метаболизме растений и уменьшением способности фиксировать молекулярный азот в бобовых растениях. Особенно интенсивно поглощение тяжелых металлов растениями происходит на кислых грунтах. Причем наиболее опасными по степени загрязнения почв тяжелыми металлами являются почвы многогумусовые, глинистосуглинистые, темно-серые лесные, черноземы и темно-каштановые [Lockhart H., Blakeley R. //Environ Sci. Tech. – 1975. – V. 9.– P. 1035].
Другая опасность состоит в том, что тяжелые металлы, связанные ЭДТА, образуя новые комплексонаты, не только усваиваются растением, но и попадают в грунтовые воды. Это приводит к накоплению биометаллов в высоких концентрациях, что ведет к разрушению экосистем. Примером подобной экологической катастрофы, произошедшей в результате неконтролируемого применения ЭДТА, может служить дельта реки Миссисипи в США, в которой сейчас извергается поток, обогащенный ЭДТА и другими химическими веществами, способствующими увеличению ежегодной «мертвой зоны», охватывающей огромную территорию [Автухович И. Е.,Постников Д. А. ВлияниеЭДТА на поведение металлов в субстрате и их накопление растениями //Естественные и технические науки. – 2014].
То, что попадание продуктов, содержащих ЭДТА, в окружающую среду создает значительные экологические проблемы также подтверждается результатами исследований ряда зарубежных ученых [Nowack B., VanBriesen J.M. Chelating agents in the environment //Biogeochemistry of Chelating Agents/eds. B. Nowack, J. M. VanBriesen / ACS Symposium Series 910 – N.Y.: Washington, DC, 2005 – P. 1–18 и т.д.].
Читайте также: Модели юбок из тонких тканей
В Канаде и многих странах Европы ЭДТА запрещена из-за своей токсичности.
В научной литературе отмечается и токсичность самой ЭДТА для некоторых видов растений. Так препарат ЭДТА в дозе 6 ммоль/кг грунта оказался токсичным для сафлора красильного (Carthamus tinctorius), что проявилось в виде хлороза листьев и снижения биомассы растений [Автухович И.Е., Постников Д.А. Ремедиация грунтов. Индуцированная витоэкстракция. / Saabibcken, Germany: Palmarium academic publishing. – 2013. – 92 c.].
Являясь относительно малотоксичной для человека, ЭДТА проявляет цитотоксичность и генотоксичность для животных. Острую токсичность с летальным исходом вызывает у крыс доза,равная 2-2,2 г/кг живого веса [Gardiner J. Complexation of trace metals by ethylenediaminetetraacetic acid (EDTA) in natural waters // Water Res. – 1976 – Vol. 10 – P. 507–514].
На основании вышеизложенного можно сделать вывод, что использование ЭДТА в сельском хозяйстве для внесения микроэлементов сопряжено с определенным риском, уровень которого повышается с увеличением концентрации данного хелатанта в почве.
Неконтролируемое применение комплексонов на основе ЭДТА может ухудшить экологическую обстановку в регионе, негативно влиять на потребительские свойства сельскохозяйственной продукции и последующую урожайность сельскохозяйственных культур.

Юрий Самусенко,
кандидат химических наук
Ответы на задачи для Коллоквиума № 1
№ 1 Известно, что ионные каналы мембраны возбудимой клетки регулируют амплитуду мембранных потенциалов. Экспериментально было обнаружено, что яд тетродотоксин блокирует натриевые каналы мембраны возбудимой клетки.
Как изменится при этом потенциал покоя и потенциал действия нервного волокна? Повлияет ли тетродотоксии на проведение возбуждения по нервному волокну?
1) Потенциал покоя создаётся преимущественно за счёт выхода ионов калия по концентрационному градиенту из клетки. При этом натриевые каналы частично открыты, и некоторое количество ионов натрия проходит в клетку, уменьшая потенциал покоя. Следовательно, блокада натриевых каналов тетродоксином приведёт к небольшому увеличению потенциала покоя.
2) Поскольку проницаемость натриевых каналов увеличивается при возникновении потенциала действия, их блокада тетродоксином приведёт к невозможности распространения возбуждения по нервному волокну.
№ 2 Известно, что градиент концентрации натрия между двумя сторонами клеточной мембраны влияет на величину потенциалов покоя и действия возбудимой клетки. В эксперименте увеличили концентрацию ионов натрия внутри нервной клетки вначале весьма незначительно, затем выровняли концентрацию ионов натрия внутри клетки и в окружающей клетку среде.
Как изменится величина потенциалов покоя и действия в двух описанных ситуациях?
1) Небольшое увеличение концентрации положительно заряженных ионов натрия внутри клетки уменьшит разность потенциалов снаружи и внутри клетки, что вызовет небольшую деполяризацию. Разность потенциалов приблизится к критическому уровню деполяризации, следовательно, возбудимость повысится, и потенциалы действия будут возникать легче.
2) При выравнивании концентрации ионов натрия внутри клетки и снаружи клетки потенциал действия не сможет возникнуть, поскольку натрий не сможет входить в клетку в условиях отсутствия градиента концентрации.
№ 3 Известно, что фазы потенциала действия нервного волокна (быстрая деполяризация и реполяризация) возникают вследствие движения ионов натрия и калия вдоль концентрационных градиентов и не требуют непосредственной затраты энергии. В эксперименте на нервное волокно подействовали уабаином — веществом, подавляющим активность АТФ-азы.
Изменится ли величина потенциала покоя и потенциала действия в обработанном уабоином нервном волокне? Изменится ли проведение возбуждения по обработанному уабаином нервному волокну?
Потенциал покоя и потенциал действия возбудимых клеток обусловлены разной концентрацией ионов, в первую очередь, калия и натрия, снаружи и внутри клетки. Разность концентраций ионов поддерживается благодаря калиево-натриевому насосу, работая которого является энергозависимой и требует АТФ-азной активности. Следовательно, ингибирование АТФ-азы приведёт к выравниванию концентраций калия и натрия снаружи и внутри клетки. Это приведёт к 1) исчезновению потенциала покоя и 2) полной невозможности возникновения и проведения потенциала действия.
№ 4 Известно, что препарат ЭДТА связывает в живых тканях ионы кальция. В экспериментальных условиях на нервно-мышечном препарате лягушки в область нервно-мышечного синапса ввели ЭДТА.
Как изменится процесс проведения возбуждения в синапсе? Повлияет ли ЭДТА на синтез ацетилхолина, проницаемость пресинаптической мембраны для медиатора, генерацию постсинаптического потенциала, активность холинэстеразы?
При проведении возбуждения в пресинаптическую область нервного волокна увеличивается проницаемость поверхностной мембраны, и ионы кальция входят по градиенту концентрации внутрь волокна. Связываясь с везикулами кальций обеспечивает движение везикул в сторону пресинаптической мембраны, что необходимо для высвобождения медиатора в синаптическую щель.
1) Связывание ионов кальция ЭДТА приведёт к прекращению высвобождения медиатора в нервно-мышечном синапсе и блокаде проведения через синапс.
2) ЭДТА не повлияет на синтез ацетилхолина. ЭДТА не повлияет непосредственно на проницаемость пресинаптической мембраны для медиатора и возможность генерации постсинаптического потенциала. Однако эти процессы не будут происходить без медиатора. Активность холинэстеразы не изменится, но без медиатора она не будет проявляться.
№ 5 Известно, что проведение возбуждения в синапсе состоит из нескольких стадий. В эксперименте воздействие химического вещества на нервно-мышечные синапсы привело к прекращению передачи возбуждения с нерва на скелетную мышцу. При введении в указанную область ацетилхолина проведение возбуждения через синапс не восстановилось. Введение фермента ацетилхолинэстеразы восстановило проведение возбуждения.
Какие могут быть механизмы прекращения проведения возбуждения в синапсе? Как эти виды блокады проведения возбуждения связаны с последовательными процессами проведения возбуждения в синапсе? Каков механизм действия изучаемого вещества на нервно-мышечный синапс?
1) Возможными причинами прекращения перехода возбуждения с нерва на мышцу в синапсе могут быть: нарушение выделение медиатора ацетилхолина пресинаптической областью; инактивация или блокада холинэргических рецепторов постсинаптической мембраны; ингибирование фермента, разрушающего медиатор.
Читайте также: Ткань galaxy 06 велюр серый
2) При добавлении ацетилхолина нервно-мышечная передача не восстановилась, следовательно, дело не в недостаточном выделении медиатора. Добавление фермента холинэстеразы, расщепляющего ацетилхолин и освобождающего рецепторы постсинаптической мембраны для взаимодействия со следующими квантами медиатора, восстановило синаптическую передачу. Следовательно, изучаемый препарат является ингибитором холинэстеразы.
№ 6 Известно, что гладкие мышцы имеют ряд физиологических особенностей по сравнению со свойствами скелетных мышц. В ходе эксперимента из стенки кишечника и стенки артерии мышечного типа животного было выделено по фрагменту (длиной 2 см и шириной 2 мм), содержащему гладкомышечные волокна. Третий фрагмент такого же размера был выделен из скелетной мышцы. Внешне мало отличающиеся друг от друга фрагменты поместили в камеру с физиологическим раствором, что обеспечивало условия для их жизнедеятельности в течение некоторого времени.
Как различить принадлежность фрагментов мышечной ткани по их функциональным свойствам? По какому функциональному признаку, без применения воздействий, можно идентифицировать принадлежность одного из фрагментов к мышечной ткани кишечника? Как с помощью раздражения фрагментов мышечной ткани можно отличить мышечную ткань внутренних органов от скелетной мышцы?
1) При наличии морфологического сходства между тремя фрагментами мышечной ткани, фрагменты гладкомышечной ткани отличаются автоматизмом, то есть способностью к спонтанной генерации потенциалов действия и сокращению.
2) Автоматизм хорошо выражен у гладких мышц стенок полых органов, в частности, кишечника, и не характерна для гладких мышц стенок кровеносных сосудов.
3) Гладкие мышцы обладают высокой чувствительностью к биологически активным веществам в отличие от скелетных мышц. Подведение раствора ацетилхолина вызовет ритмические сокращения фрагмента кишечника. Раствор адреналина вызовет спастическое сокращение фрагмента артерии. Действие химических веществ не вызовет сокращение фрагмента скелетной мышцы. Сокращение фрагмента скелетной мышцы можно вызвать электрическим раздражением.
№ 7 Известно, что возбудимость является одним из основных свойств возбудимых тканей. Экспериментально сравнивали возбудимость нервной и мышечной ткани до и после длительного прямого и непрямого раздражения мышцы. Было установлено, что исходно возбудимость одной ткани больше чем второй. Кроме того, было зафиксировано изменение возбудимости после длительного раздражения.
Как был поставлен эксперимент для сравнения возбудимости? Какая ткань и почему имела большую возбудимость? Как изменилась возбудимость нерва и мышцы после длительного прямого и непрямого раздражения мышцы?
1) Эксперимент проводили на нервно-мышечном препарате лягушки. Сначала нерв, затем мышцу раздражали одиночными электрическими импульсами. Силу раздражения постепенно увеличивали до появления порога мышечного сокращения. Таким образом, определяли порог раздражения нерва и мышцы.
2) Нервная ткань имела большую возбудимость, так как её порог раздражения был меньше, чем мышечной ткани.
3) Возбудимость понизится в случае уменьшения концентрационных градиентов на клеточной мембране при длительном раздражении.
№ 8 Известно, что возбудимость является одним из основных свойств возбудимых тканей. Экспериментально было установлено, что после нанесения на изолированный нерв и полоску миокарда надпорогового раздражения в нервном волокне и миокардиальном волокне возникали потенциалы действия, в ходе которых возбудимость то увеличивалась, то уменьшалась и иногда исчезала.
Как регистрировали электрические процессы в нерве и миокарде, возникающие после нанесения раздражения? Как регистрировали изменения возбудимости? Какая природа связи между отдельными фазами потенциала действия и изменением возбудимости?
1) Биопотенциалы регистрировали с помощью микроэлектродной техники.
2) Изменения возбудимости регистрировали с помощью измерений порога раздражения в различные фазы потенциала действия.
3) Для анализа связи необходимо нарисовать потенциалы действия и синхронные изменения возбудимости. Изменения возбудимости обратно пропорциональны изменению порога раздражения.
№ 9 Известно, что вокруг клеточных мембран возбудимых тканей существует неравномерное распределение ионов. Экспериментально увеличивали градиент концентрации снаружи и внутри возбудимой клетки раздельно для ионов Na, К, Сl и Са.
Как изменится величина потенциалов покоя и действия при увеличении градиента концентрации раздельно для ионов Na, К, Сl и Са?
1) При увеличении градиента концентрации Na величина потенциал покоя понизится, величина потенциала действия повысится.
2) При увеличении градиента концентрации К величина потенциал покоя повысится, величина потенциала действия не изменится.
3) При увеличении градиента концентрации Cl величина потенциал покоя повысится, величина потенциала действия не изменится.
4) При увеличении градиента концентрации Са величина потенциал покоя понизится, величина потенциала действия не изменится.
№ 10 Известно, что процесс возбуждения нервной и мышечной ткани характеризуется изменением ионной проницаемости клеточной мембраны. Экспериментально проводили избирательную блокаду мембранной проницаемости для ионов Na, К, Cl и Са в нервных волокнах и мышечных волокнах скелетной, гладкой и миокардиальной мышц.
Как после этого изменится величина потенциалов покоя и действия в этих волокнах?
1) При блокаде мембранной проницаемости для Na величина потенциала покоя повысится, величина потенциала действия понизится.
2) При блокаде мембранной проницаемости для K величина потенциала покоя понизится, величина потенциала действия не изменится.
3) При блокаде мембранной проницаемости для K и для Cl величина потенциала покоя понизится, величина потенциала действия не изменится.
4) При блокаде мембранной проницаемости для K и для Ca величина потенциала покоя повысится, величина потенциала действия не изменится. В миокардиальных и гладкомышечных волокнах длительность потенциалов действия уменьшится в связи с ускорением реполяризации.
№ 11 Известно, что суммация одиночных сокращений является базисным свойством мышечной ткани. Экспериментально было установлено, что скелетная, гладкая и миокардиальная мышцы обладают разной способностью к суммации.
Читайте также: Паплин или поплин ткань состав
Какие условия надо выполнить, чтобы добиться суммации одиночных сокращений в каждом виде мышечной ткани в эксперименте?
1) Для суммации одиночных сокращений необходимо соблюдение двух условий. Второе раздражение должно наноситься в течение первого одиночного сокращения и не должно попасть период рефрактерности первого возбуждения.
2) Длительность возбуждения и рефрактерного периода в скелетной мышце соответствует латентному периоду ее одиночного сокращения. В гладкой мышце длительность возбуждения и рефрактерный период соответствует латентному периоду и фазе укорочения. В миокардиальной мышце длительность возбуждения и рефрактерный период соответствует длительности одиночного сокращения.
3) Поэтому, скелетная мышца способна к слитному титаническому сокращению. Гладкая мышца также способна к суммации одиночных сокращений при меньшей частоте раздражения, что связанно с ее меньшей лабильностью. Миокардиальная мышца не способна к суммации одиночных сокращений в условиях нормы. Однако в условиях патологии в сердце может возникнуть титаническое сокращение, что связано с укорочением периода рефрактерности.
№ 12 Известно, что лабильность или функциональная подвижность является важной характеристикой возбудимых тканей. Экспериментально, была исследована лабильность возбудимых тканей лягушки: миелиновых волокон седалищного нерва, безмиелиновых волокон одного из симпатических нервов, нервно-мышечных синапсов и мышечных волокон икроножной мышцы, желудка и сердца.
Как определяли лабильность у различных возбудимых образований? Какой параметр обычно используют как меру лабильности? У каких исследованных образований лабильность выше или ниже и почему? Как лабильность может характеризовать возбудимость исследуемых образований?
1) Лабильность определяют с помощью нанесения ритмического раздражения с увеличивающейся частотой и регистрацией процессов возбуждения или сокращения исследуемой возбудимой ткани.
2) Мерой лабильности является максимальная частота раздражения, которую возбудимая ткань может воспроизвести без трансформации ритма.
3) Мера лабильности обратно пропорциональна длительности рефрактерного периода. Миелиновые нервные волокна обладают наибольшей лабильностью. Безмиелиновые нервные волокна, мышечные волокна скелетных мышц, нервно-мышечные синапсы и кардиомиоциты имеют меньшую лабильностью. Гладкие мышцы внутренних органов обладают наименьшей лабильностью.
4) Лабильность является одной из косвенных характеристик возбудимости. Чем больше лабильность, тем больше возбудимость.
№ 13 Известно, что поддержание постоянства ионного состава в возбудимых тканях необходимо для их нормального функционирования. Отсутствие солей и, в частности, солей кальция в питьевой воде населения отдаленного района России привело к нарушению функций мышечной ткани. Обследование населения показало, что даже практически здоровые люди жаловались на повышенную мышечную утомляемость и недостаточную физическую силу.
Какая роль ионов кальция в механизме мышечного сокращения? Как снижение концентрации кальция скажется на работе сердца? Какие обследования целесообразности для изучения функций скелетной мускулатуры и миокарда?
1) Выходящий из Т систем и эндоплазматического ретикулума Са соединяется с тропонином, в результате чего головки выростов миозина вступают в контакт с актином, обеспечивая скольжение актина вдоль миозина и уменьшение длины сакромеров.
2) Уменьшение концентрации Са вызовет отрицательный инотропный эффект в сердце. Одновременно фаза реполяризации станет короче, что уменьшит рефрактерный период. Это может привести к нежелательной для работы сердца суммации одиночных сокращений.
3) Для изучения силы и выносливости скелетной мускулатуры необходимо исследовать динамометрию. Для изучения выносливости сердца необходимо провести исследование функций сердца с физическими нагрузками.
№ 14 Известно, что блокатор синаптической передачи возбуждения может вызвать разные изменения в скелетных мышцах и мышцах внутренних органов. В ходе обследования испытуемого было установлено, что блокатор ацетилхолинергической передачи возбуждения в синапсах атропин вызвал расширение зрачка, увеличение частоты и силы сердечных сокращений, уменьшение перистальтики желудочно-кишечного тракта. При этом не изменилась сократительная функция скелетной мускулатуры.
Объясните возможные причины различного действия блокаторов холинэргической синаптической передачи возбуждения в нервно-мышечных синапсах соматической нервной системы и в синапсах вегетативной нервной системы на внутренних органах.
1) Медиатор ацетилхолин действует на два вида постсинаптических рецепторов: М- и Н- холинорецепторы. М- холинорецепторы находятся в нейроорганных синапсах и вегетативных ганглиях.
2) Атропин блокирует только М-холинорецепторы. Поэтому блокируется действие парасимпатической нервной системы, а соматическая регуляция скелетной мускулатуры не нарушается.
№ 15 Известно, что суммарный потенциал действия нерва складывается из потенциалов действия одиночных нервных волокон, входящих в нерв. Экспериментально исследовали суммарный потенциал действия изолированного седалищного нерва, выделенного из крупного животного. Раздражение наносили на центральный конец нерва. Ближе к месту раздражения была зарегистрирована синусоидальная форма суммарного потенциала действия. На периферическом конце нерва суммарный потенциал состоял из нескольких пиков и волн. Амплитуда его значительно уменьшилась.
С чем связано изменение формы суммарного потенциала действия по ходу проведения возбуждения в нерве? Как диаметр волокна и наличие миелина влияют на скорость проведения возбуждения? Почему происходит уменьшение амплитуды суммарного потенциала? Работает ли при этом закон «Всё или ничего»?
1) Расслоение суммарного потенциала действия на отдельные волны связного с различной скоростью проведения возбуждения в волокнах образующих нерв.
2) Скорость проведения возбуждения больше в миелиновых волокнах и волокнах с большим диаметром.
3) Амплитуда суммарного потенциала уменьшается по мере ответвления от нерва нервных волокон.
4) Закон “все или ничего” при этом не работает. Суммарный потенциал действия нерва прямо зависит от количества волокон входящих в нерв. Потенциал действия каждого волокна, входящего в нерв, подчиняется закону “все или ничего”.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом