
По существу, любой фактор, способный вызвать диффузию достаточного числа ионов натрия через мембрану внутрь клетки, может «запустить» автоматическое регенеративное открытие натриевых каналов. Это возможно в результате влияний на мембрану механических, химических или электрических раздражителей.
Все они используются в разных участках тела для генерации потенциалов действия в нервах или мышцах: механическое давление — для возбуждения чувствительных нервных окончаний в коже, химические нейромедиаторы — для передачи сигналов от одного нейрона к другому в мозге, электрический ток — для передачи сигналов между прилежащими мышечными клетками в сердце и кишечнике. Чтобы понять процесс возбуждения, обсудим принципы электрической стимуляции.
Возбуждение нервного волокна отрицательно заряженным металлическим электродом. В условиях экспериментальной лаборатории обычным способом возбуждения нерва или мышцы является электрическое раздражение через размещенные на их поверхности два небольших электрода, один из которых — отрицательный, другой — положительный. В данном случае возбудимая мембрана стимулируется отрицательным электродом.
Рассмотрим причины влияния отрицательного электрода. Вспомните, что потенциал действия инициируется открытием электроуправляемых натриевых каналов, а эти каналы, как известно, открываются при понижении нормального потенциала покоя мембраны. Ток отрицательного электрода снижает положительный потенциал на наружной стороне мембраны, приближая его к отрицательному значению потенциала внутри волокна.
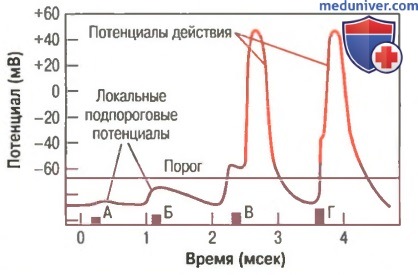
Значение увеличения силы стимула для возникновения потенциала действия. Развитие локальных подпороговых потенциалов в ответ на стимулы, сила которых ниже порогового значения, необходимого для появления потенциала действия.
Это уменьшает трансмембранную разность потенциалов и открывает натриевые каналы, в результате возникает потенциал действия. Напротив, у положительного электрода, где на наружную поверхность мембраны нерва попадают положительные заряды, трансмембранная разность потенциалов не уменьшается, а повышается. Следовательно, здесь возникает гиперполяризация — состояние, снижающее возбудимость, что не способствует развитию потенциала действия.
Порог возбуждения и локальные потенциалы. Слабый отрицательный электрический стимул может не возбудить волокно. Однако при усилении стимула наступает момент, когда возбуждение возникает. На рисунке показано влияние последовательно наносимых стимулов возрастающей силы. Очень слабый стимул в точке А изменяет мембранный потенциал с -90 до -85 мВ, но этого изменения недостаточно для развития автоматических регенеративных процессов, необходимых для возникновения потенциала действия.
В точке Б сила стимула больше, однако интенсивность его все же недостаточна. Тем не менее, каждый из этих слабых стимулов вызывает местное изменение мембранного потенциала длительностью в 1 мсек и более. Эти местные изменения потенциала называют локальными потенциалами, но если они не способны вызвать потенциал действия, их называют подпороговыми потенциалами. В точке В на рис. 5-18 стимул еще сильнее. На этот раз локальный потенциал достиг минимального уровня, необходимого для возникновения потенциала действия, называемого пороговым уровнем, но потенциал действия возникает лишь после короткого латентного периода.
В точке Г стимул еще сильнее, амплитуда локального потенциала больше, и потенциал действия возникает после более короткого латентного периода.
Следовательно, даже очень слабый стимул вызывает местные изменения мембранного потенциала, но интенсивность локального потенциала должна подняться до порогового уровня, прежде чем возникнет потенциал действия.
Видео физиология возбуждения тканей (потенциал покоя, потенциал действия) — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Что происходит в клетке при возбуждении. помогите биологию
Возбуждение — ответная реакция живой ткани на внешнее воздействие, выражающаяся изменением характера или интенсивности протекающих в ней процессов; в узком смысле — это физиологический процесс, которым некоторые типы клеток (нервные, мышечные, железистые) отвечают на внешнее воздействие (раздражитель) . Способность клеток и тканей реагировать на раздражение возбуждением называется возбудимостью. Минимальная сила раздражителя, на которую возбудимая ткань отвечает возникновением возбуждения, называется пороговой; чем меньше эта величина, тем выше уровень возбудимости и ткань легче возбуждается.
В процессе эволюции у отдельных компонентов клетки, в частности белковых структур, первоначально возникла способность восстанавливать свою структуру, поврежденную извне. Затем возникла способность избегать опасность и, наконец, высшая форма возбуждения — сигнализация, необходимая для нормального существования многоклеточного организма.
У многоклеточных организмов возбуждение стало основной функцией клеток нервной ткани. Возбуждение и обратное ему явление — торможение — основа всех видов нервной деятельности, в т. ч. психической (см. Высшая нервная деятельность) . Возбуждение мышечных и железистых клеток характеризуется переходом этих клеток от состояния покоя к состоянию физиологической деятельности, свойственной этим клеткам — сокращению для мышечных и секреции (выделение) — для железистых клеток . При возникновении процесса возбуждения в возбудимых тканях происходит усиление обмена веществ, что сопровождается тратой энергетических запасов, накопленных в клетках . При длительном возбуждении (например, в клетках нервной ткани) происходит истощение запасов энергетических (питательных) веществ, следствием чего может быть развитие явлений «нервного истощения» , сопровождающегося различными невротическими, истерическими и другими состояниями.
Различают возбуждение местное и возбуждение распространяющееся.
Местное возбуждение — это изменение электрических свойств на каком-либо отдельном участке клеточной оболочки, возникающее в результате перераспределения ионов по ее обеим сторонам. Этот вид возбуждения играет определенную роль лишь на ограниченном участке — в пределах одной клетки и неспособно вызывать возбуждение какой-либо другой, даже соседней, клетки . Вместе с тем именно местное возбуждение является фактором, «запускающим» специфическую деятельность клеток (например, изменение проницаемости клетки, выделение секрета, возникновение нервного импульса) .
Распространяющееся возбуждение является особой формой возбуждения, которое было выработано природой для компенсации неспособности местного возбуждения передаваться на большие расстояния (в пределах всего организма) . Возникнув однажды (например, в специальных нервных образованиях — рецепторах — под влиянием света, звука, тепла и т. д.) , местное возбуждение (при условии, что раздражающее воздействие достаточно сильно) становится самоподдерживающимся и начинает распространяться по клетке с постоянной скоростью. Импульсы распространяющегося возбуждения передаются в центральную нервную систему, откуда в виде ответных импульсов поступают к исполнительным органам (мышцам, сосудам, железам) , в которых через механизмы уже местного возбуждения вызывают соответствующие реакции (например, производится движение, сокращение стенок кровеносного сосуда, выделение секрета и т. д.) .
Читайте также: Мебельный хлопок что это за ткань
При возбуждении любой ткани происходит воздействие
В возбудимой ткани невозможно вызвать новый потенциал действия, пока мембрана возбужденного волокна остается деполяризованной в связи с развитием предыдущего потенциала действия. Это связано с тем, что вскоре после возникновения потенциала действия натриевые каналы (или кальциевые каналы, или оба типа каналов) инактивируются, и возбуждающий сигнал любой силы, действующий на ткань в этот момент, не может открыть инактивационные ворота. Единственным условием для их повторного открытия является возврат мембранного потенциала к исходному или близкому к исходному уровню. Затем в течение небольшой доли секунды инактивационные ворота каналов открываются, и становится возможным развитие нового потенциала действия.
Период, в течение которого нельзя вызвать второй потенциал действия даже сильным стимулом, называют абсолютным рефрактерным периодом. Для крупных миелинизированных нервных волокон этот период составляет примерно 1/2500 сек. Легко рассчитать, что такое волокно может максимально передавать около 2500 имп/сек.
Кроме факторов, повышающих возбудимость нервного волокна, существуют так называемые мембраностабилизирующие факторы, способные снижать возбудимость. Например, высокая концентрация ионов кальция во внеклеточной жидкости уменьшает проницаемость мембраны для ионов натрия, снижая возбудимость. В связи с этим ионы кальция называют стабилизатором.
Местные анестетики. К наиболее важным стабилизаторам относят многие вещества, используемые в клинике в качестве местных анестетиков, в состав которых входят прокаин и тетракаин. Большинство из них действуют непосредственно на активационные ворота натриевых каналов, затрудняя их открытие, что сопровождается снижением возбудимости мембран. После снижения возбудимости до уровня, при котором отношение амплитуды потенциала действия к порогу возбуждения (называемое фактором надежности) опускается ниже 1,0, импульсы по анестезированному нерву не проходят.
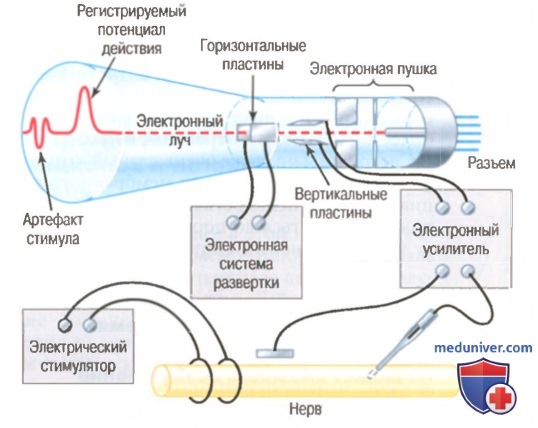
Катодно-лучевой осциллоскоп для регистрации быстрых потенциалов действия.
Катодный осциллограф. Ранее в этой главе мы отмечали, что изменения мембранного потенциала во время генерации потенциала действия происходят чрезвычайно быстро. Действительно, развитие большей части комплекса потенциала действия в крупных нервных волокнах занимает менее 1/1000 сек. На некоторых рисунках данной главы показан электроизмерительный прибор, регистрирующий эти изменения потенциалов. Однако ясно, что реакции любого прибора, способного регистрировать потенциалы действия, должны быть чрезвычайно быстрыми. Для практических целей единственным широко используемым прибором, способным точно реагировать на быстрые изменения мембранного потенциала, является катодно-лучевой осциллоскоп.
На рисунке показаны основные компоненты катодно-лучевого осциллоскопа. Катодно-лучевая трубка состоит из электронной пушки и флюоресцентного экрана, который «бомбардируется» электронами. При ударе электронов о поверхность экрана флюоресцентный материал светится. Если луч электронов перемещается по экрану, вместе с ним движется яркое пятно света, оставляя на экране флюоресцирующую линию.
Кроме электронной пушки и флюоресцентной поверхности, катодно-лучевая трубка снабжена двумя парами электрически заряженных пластин. Одна пара расположена с обеих сторон от электронного луча, а другая — сверху и снизу. Соответствующие электронные усилители изменяют напряжение на этих пластинах таким образом, что электронный луч отклоняется вверх или вниз в ответ на электрические сигналы, приходящие от регистрирующих электродов. Под влиянием внутреннего электронного блока осциллоскопа луч электронов перемещается по экрану горизонтально с постоянной скоростью. При этом регистрируется кривая, которую видно на экране катодно-лучевой трубки рисунка, с временной разверткой по горизонтали и изменениями потенциалов, регистрируемых отводящими электродами, по вертикали.
На левом конце кривой виден небольшой артефакт стимула, связанный с электрическим стимулом, который используется для вызова потенциала действия, справа на кривой — сам потенциал действия.
Видео физиология возбуждения тканей (потенциал покоя, потенциал действия) — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Кафедра физиологии
Разделы сайта
Новости
Общие представления о физиологии возбудимых тканей
Общие представления о физиологии возбудимых тканей
Биологическая реакция – ответная реакция клеток, тканей, органов в ответ на раздражитель (стимул).
Раздражимость – свойство всех живых тканей изменять своё внутреннее состояние при изменении внешних условий.
Виды тканей в зависимости от реагирования на внешние раздражители:
I Возбудимые — обладают свойством возбудимости, т.е. способностью возбуждаться нервная, мышечная, железистая.
II Невозбудимые – изменяют свое состояние, но не генерируют процесс возбуждения в ответ на нанесенный стимул.
Возбудимость – способность ткани переходить в возбуждённое состояние.
Возбуждение – деятельное состояние тканей в ответ на действие раздражителя, это сложная биологическая реакция, проявляющаяся в совокупности физических, физико-химических и функциональных изменений, способная к распространению по ткани.
Возбуждение включает в себя неспецифические и специфические компоненты.
Неспецифические:
сдвиг химических реакций, образование тепла, физико-химические изменения,
продукция биопотенциалов, структурные изменения в мембране клеток.
Специфические:
мышечная ткань отвечает мышечным сокращением, нервная ткань — генерацией нервного импульса и его проведением, железистая ткань – образованием и выделением секрета.
Возбуждение может быть локальным и динамическим (распространяющимся).
Биопотенциалы
Луиджи Гальвани 1791 г. в эксперименте показал, что живые ткани содержат «животное электричество», его научный оппонент, физик Вольта — что это электричество от разнородных металлов, он создал первый источник постоянного тока, который носит название гальванический элемент.
Виды биопотенциалов:
1. Биопотенциал покоя (мембранный) — МПП.
2. Биопотенциал действия (возбуждения) — ПД.
- Биопотенциал покоя – это разность потенциалов между наружной и внутренней поверхностью мембраны клетки в покое. Наружная поверхность мембраны клетки имеет положительный заряд, а внутренняя – отрицательный.
Биопотенциал покоя регистрируется внутриклеточным методом – с помощью микроэлектродов, один из которых вводится внутрь клетки (рис.1).
Рисунок 1. Схематическое представление метода регистрации биопотенциалов.
В эксперименте биопотенциал покоя можно зарегистрировать между повреждённым и неповреждённым участком ткани. Повреждённый участок является моделью внутренней поверхности мембраны клетки.
При внутриклеточном отведении перезарядка мембраны регистрируется под одним электродом (однофазный ПД), при внеклеточном отведении потенциал действия проходит через два электрода (регистрируется двухфазный ПД).
- Биопотенциал действия – это кратковременные высокоамплитудные изменения МПП, которые возникают при возбуждении. ПД регистрируется в раздражаемых тканях, в которых возникает волна возбуждения (рис.2). Измеряется ПД с помощью внутриклеточного отведения и внеклеточного отведения.
Читайте также: Обшить дверь тканью в машине

Рисунок 2. Потенциал действия, основные его фазы.
Современная, экспериментально доказанная, мембранно-ионная теория возникновения биопотенциалов (Ходжкин, Хаксли, Катц).
Основные положения:
- Электрические процессы возникают на плазматической мембране клетки, которая состоит из бимолекулярного слоя липидов (остов мембраны) и белков, которые выполняют различные функции в мембране: рецепторную, ферментативную, образуют в ней каналы и насосы (рис.3).
Канал мембраны может быть неспецифическим, он постоянно открыт, не имеет воротного механизма, электрические воздействия не изменяют его состояния. Называют каналом «утечки». Специфические каналы (селективные) имеют воротный механизм, поэтому могут находиться или в открытом, или в закрытом состоянии в зависимости от электрических воздействий на мембрану и пропускают только определенный ион. Этот канал состоит из трех частей: водной поры – выстлана внутри гидрофильными группами; селективного фильтра – на наружной поверхности, который пропускает ионы в зависимости от их размера и формы; ворот – на внутренней поверхности мембраны, управляют проницаемостью канала.

Рисунок 3. Строение биологической мембраны.
Канал мембраны может быть неспецифическим, он постоянно открыт, не имеет воротного механизма, электрические воздействия не изменяют его состояния. Называют каналом «утечки». Специфические каналы (селективные) имеют воротный механизм, поэтому могут находиться или в открытом, или в закрытом состоянии в зависимости от электрических воздействий на мембрану и пропускают только определенный ион. Этот канал состоит из трех частей: водной поры – выстлана внутри гидрофильными группами; селективного фильтра – на наружной поверхности, который пропускает ионы в зависимости от их размера и формы; ворот – на внутренней поверхности мембраны, управляют проницаемостью канала (рис.4).

Рисунок 4. Строение ионного канала.
Каналы для натрия имеют два типа ворот: быстрые активационные и медленные инактивационные. В покое открыты медленные инактивационные и закрыты быстрые активационные. При возбуждении происходит открытие быстрых активационных и медленное закрытие медленных инактивационных, т.е. на короткий промежуток времени оба типа ворот открыты (рис.5).

Рисунок 5. Работа активационные и инактивационный воротных механизмов натриевого ионного канала.
Калиевые каналы имеют только медленные ворота.
Насосы выполняют функцию транспорта через мембрану ионов против градиента концентрации, для их работы используется энергия АТФ.
- По обе стороны мембраны существует концентрационный градиент.
Внутри клетки в 40 раз > К+; t;/p>
Вне клетки: в 20-30 раз > Na+,
в 50 раз > Cl-.
- Мембрана пропускает молекулы жирорастворимых веществ, а анионы органических кислот не проходят. Мембрана проницаема для воды, для ионов проницаемость мембраны различна: для калия в состоянии покоя проницаемость почти в 25 раз больше, чем для натрия. При возбуждении увеличивается проницаемость и для калия (постепенно), и для натрия (быстро, но на очень короткий промежуток времени).
Потенциал покоя
Проницаемость мембраны для ионов К+ повышена, поэтому калий играет основную роль в генерации МПП. Калий создаёт электрическое поле и заряжает наружную поверхность мембраны «+». В тот момент, когда «+» потенциал наружной стороны достигает определённой величины по отношению к «–» внутри, который создается анионами – наступает динамическое равновесие между входящими и выходящими из клетки ионами К+. Этому моменту соответствует потенциал равновесия для К — потенциал покоя.
МПП характеризуется:
1. постоянством;
2. полярностью, снаружи «+», внутри «-»;
3. величина – в мВ, для скелетной мышцы — 60 – 90 мВ,
для гладкой — -30 – 70 мВ,
для нерва -50 – 80мВ,
для секреторной клетки — -20мВ.
МПП — один из основных показателей состояния физиологического покоя клетки. При увеличении внеклеточной концентрации калия уменьшается МПП, т.к. уменьшается диффузия калия из клетки в связи со снижением его концентрационного градиента. При действии веществ, блокирующих ресинтез АТФ, т.к. прекращается работа натрий-калиевого насоса, также снижается МПП. Ионы натрия и хлора входят в клетку, но ввиду низкой проницаемости значительного влияния на МП не оказывают.
Потенциал действия
При возбуждении – резко увеличивается (в несколько тысяч раз) проницаемость для ионов Na, которые поступают внутрь клетки лавинообразно и заряжают внутреннюю сторону «+» — происходит деполяризация мембраны, а затем количество ионов натрия внутри превышает калиевый заряд на поверхности и это приводит к перезарядке мембраны (реверсии). Постепенно увеличивающаяся проницаемость для калия и его поток из клетки инактивирует натриевую проницаемость и приводит к восстановлению заряда на мембране. Возникает фаза реполяризации.
Существенным фактором является натрий-калиевый насос, который выводит из клетки 3 иона натрия в обмен на 2 иона калия, вводимые в клетку. Его работа зависит от метаболизма клетки, в частности, от ее энергоснабжения. При этом расходуется 1 молекула АТФ (рис.6).
Рисунок 6. Механизм работы натрий-калиевого насоса.
ПД состоит из пикового потенциала, который образуется фазой деполяризации, реверсии и реполяризации, и следовых потенциалов (рис.2).
Следовые потенциалы:
Отрицательный (следовая деполяризация);
Положительный (следовая гиперполяризация).
Причиной следовых потенциалов являются дальнейшие изменения соотношения между входом натрия в клетку и выходом калия из нее. При следовой деполяризации отмечается остаточный ток натрия в клетку при одновременном снижении калиевого тока. При следовой гиперполяризации – остаточное усиление тока калия из клетки при одновременной активации натрий-калиевого насоса.
ПД характеризуется:
1. изменяющимся характером;
2. кратковременностью – несколько мсек;
3. зарядом мембраны, снаружи – «-», внутри – «+».
При действии веществ, блокирующих натриевые каналы, ПД не генерируется, т.к. в норме деполяризация мембраны обусловлена повышение ее натриевой проницаемости. При увеличении силы раздражителя выше порога амплитуда ПД не изменяется, т.к. не изменяется число активированных натриевых каналов, которые максимально раскрываются при пороговом раздражении.
Читайте также: Красивые модели из ткани
Условия, необходимые для возникновения возбуждения (законы раздражения).
Возбудимость тканей различна. Чтобы вызвать возбуждение, раздражитель должен обладать:
1. Достаточной силой – закон порога.
2. Крутизной (градиентом) нарастания этой силы – закон аккомодации.
3. Временем действия – закон силы-времени.
1. Закон силы. Мерой возбудимости является порог раздражения – минимальная сила раздражителя, способная вызвать возбуждение. Все раздражители можно разделить на подпороговые, пороговые и сверхпороговые. По биологическому значению раздражители делят на адекватные (действующие на ткань в естественных условиях, к ним она приспособлена в процессе эволюции) и неадекватные. В физиологических экспериментах в качестве раздражителя чаще всего используется электрический ток, т.к. он вызывает обратимые изменения, легко дозируется по силе и длительности, по своей природе близок к электрическим процессам, протекающим в живых организмах.
В 1870 г. Боудич в эксперименте на мышце сердца путем нанесения на неё одиночных пороговых раздражений регистрировал ответную реакцию — установил, что на подпороговое раздражение реакции не было, при пороговой силе и сверхпороговой амплитуда ответной реакции была одинаковой. На основании этого он предложил закон «Всё или ничего».
После введения в экспериментальные исследования микроэлектронной техники было установлено, что на подпороговое раздражение в ткани возникает ответная реакция. Если сила стимула меньше 50% пороговой величины, то под полюсами электродов происходит пассивная деполяризация без изменения проницаемости мембраны для ионов (электротонические изменения). Если сила стимула меньше пороговой величины, но больше 50% от нее, то в ткани возникает локальный ответ, который сопровождается деполяризацией мембраны в области нанесения раздражения и не распространяется на всю ткань, возбудимость тканей в этом участке повышена. Локальный ответ подчиняется закону силовых отношений, т.е. чем больше сила подпорогового стимула, тем больше амплитуда локального ответа. Проницаемость мембраны клетки в этом участке повышается для ионов натрия. При нанесении порогового стимула возникает ПД, амплитуда которого не изменяется, если величина стимула будет превышать пороговую, т.е. отвечает закону «Все или ничего», но на сверхпороговые стимулы длительность ПД будет меньше за счет укорочения продолжительности локального ответа.
Момент перехода локального ответа в ПД называется критическим уровнем деполяризации (КУД), а сдвиг заряда мембраны с мембранного потенциала до КУД, называется пороговым потенциалом, он наряду с порогом раздражения характеризует возбудимость ткани.
Изменение возбудимости тканей при возбуждении.
При возбуждении возбудимость тканей претерпевает определенные изменения в зависимости от фаз ПД (рис.7):
I – супернормальная возбудимость (первичная) соответствует локальному ответу, при этом два подпороговых стимула, нанесенных с интервалом времени, короче длительности локального ответа могут суммироваться и вызывать ПД;
II – абсолютная рефрактерность – соответствует регенеративной деполяризации и реверсии, при этом ткань становится абсолютно невозбудимой и не отвечает на самые сильные раздражители;
III – относительная рефрактерная фаза, соответствует реполяризации, при этом возбудимость ткани постепенно восстанавливается и сверхпороговый стимул, нанесенный в этот период может генерировать ПД;
IV – супернормальная возбудимость (вторичная или экзальфационная фаза) — следовой деполяризации, ткань становится более возбудимой, чем в исходном состоянии и даже подпороговый стимул способен вызвать ПД;
V – субнормальная возбудимость – следовой гиперполяризации, возбудимость ткани несколько снижена.

Рисунок 7. Изменение возбудимости мембраны при развитии потенциала действия.
2. Закон градиента раздражения (Дюбуа Реймон). Чем больше градиент раздражения, тем больше (до известных пределов) реакция живого образования.
За время действия медленно нарастающего стимула наступает приспособление ткани – аккомодация. Она связана с тем, что при возбуждении проницаемость для ионов натрия увеличивается на короткий промежуток времени, если в течение его раздражитель не достигает пороговой величины, то увеличивающаяся проницаемость для ионов калия инактивирует натриевую проницаемость и возбуждение не наступает. При этом происходит также сдвиг КУД с увеличением порогового потенциала.
3. Закон силы-времени (Лапик). Пороговая величина любого раздражителя находится в обратной зависимости от времени его действия, которая характеризуется математической кривой – гиперболой. Характер кривой свидетельствует о том, что подпороговые стимулы (меньше 1 реобазы) не вызовут возбуждение как долго бы они не действовали, в то же время очень сильный кратковременный стимул, длительность которого меньше полезного времени, также не вызовет возбуждение.
Сила постоянного тока, которая, действуя неопределенное время, вызывает возбуждение, называется реобазой.
Время, в течение которого ток в 1 реобазу вызывает возбуждение – полезное время.
Минимальное время, в течение которого ток силой в 2 реобазы вызовет возбуждение, называется хронаксией. Исследование этого показателя используется в неврологической и травматологической практике для изучения динамики восстановления в нервной или мышечной ткани после травмы.
Список использованной литературы
- Нормальна фізіологія /Під ред. В.І. Філімонова. – К. – Здоров’я, 1994. – С. 5 — 37.
- Физиология человека /Под ред. Г. И. Косицкого. – М., Медицина, 1985. – С. 19 – 84.
- Посібник з нормальної фізіології /Під ред. В.Г. Шевчука. – К., Здоров’я, 1995. – С. 6 — 36.
- Руководство к практическим занятиям по физиологии /Под ред. Г. И. Косицкого. – М., Медицина, 1988. – С. 72 — 94.
- Нормальная физиология /Под ред. В. И. Филимонова. — Запорожье, 1995. – С. 74-72.
- Физиология человека. Т.1 /Под ред. Р. Шмидта и Г. Тевса. – М., Мир, — 1996. — С. 9-87.
- Физиология человека. Т.1 / Под ред. В.М. Покровского. – М., Медицина, 1998. – С. 27-97.
- Общий курс физиологии человека и животных. Т.1. /Под ред. А.Д. Ноздрачева – М., Высшая школа, 1991.- С.36-116.
- Физиология человека. /Под ред. В.М. Смирнова – М., Медицина, 2002. – С. 45-61, 82-94.
- Фізіологія людини. Вільям Ф. Ганонг. – БаК, Львів, 2002. – С. 6 – 69, 74-76.