
Продолжительность потенциала действия для нервных тканей составляет
Миелинизированные и немиелинизированные нервные волокна. На рисунке показан поперечный срез типичного небольшого нерва, на котором видно много крупных нервных волокон, составляющих большую часть среза. Однако при более внимательном рассмотрении между крупными волокнами можно обнаружить множество очень мелких волокон. Крупные волокна миелинизированы, мелкие — немиелинизированы. В среднем нервный ствол содержит вдвое больше не-миелинизированных (безмякотных) нервных волокон, чем миелинизированных (мякотных).
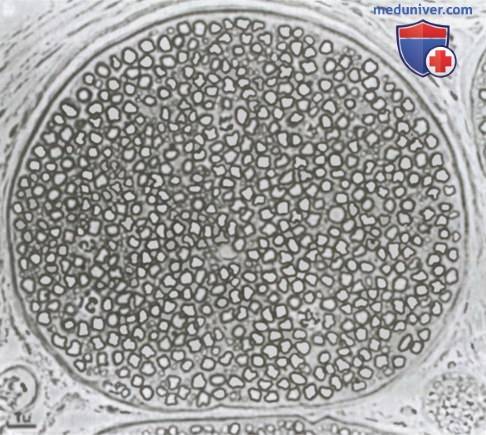 Поперечное сечение небольшого нервного ствола, содержащего миелинизированные и немиелинизированные волокна.
Поперечное сечение небольшого нервного ствола, содержащего миелинизированные и немиелинизированные волокна. 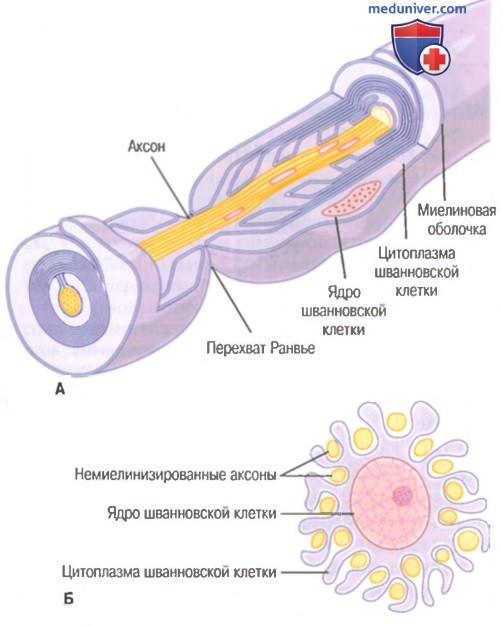 Значение шванновских клеток для изоляции нервных волокон.
Значение шванновских клеток для изоляции нервных волокон.
А. Накручивание мембраны шванновской клетки на крупный аксон формирует миелиновую оболочку миелинизированного нервного волокна.
Б. Частичная закрутка мембраны и цитоплазмы шванновской клетки вокруг множества немиелинизированных нервных волокон (показано в поперечном сечении).
На рисунке показано типичное миелинизированное волокно. Центральной его частью является аксон, по мембране которого проводится потенциал действия. Аксон заполнен аксоплазмой — вязкой внутриклеточной жидкостью. Аксон окружен миелиновой оболочкой, которая часто много толще, чем сам аксон. Примерно через каждые 1-3 мм вдоль миелиновой оболочки имеется перехват Ранвье.
Миелиновая оболочка формируется вокруг аксона шванновскими клетками. Мембрана шванновской клетки сначала охватывает аксон, затем шванновская клетка многократно вращается вокруг аксона, образуя многочисленные мембранные слои, содержащие липидное вещество сфингомиелин. Это вещество является отличным изолятором и снижает ионный ток через мембрану аксона примерно в 5000 раз. Между каждыми двумя последовательно расположенными шванновскими клетками по ходу аксона остается маленькая неизолированная область длиной всего 2-3 мкм, где ионы могут свободно переходить через мембрану аксона из внеклеточной жидкости во внутриклеточную и обратно. Эту область называют перехватом Ранвье.
Сальтаторное проведение в миелиновых волокнах от перехвата к перехвату. Ионы практически не могут проходить через толстую миелиновую оболочку мякотных волокон, однако они легко диффундируют через перехваты Ранвье. Следовательно, потенциалы действия возникают только в перехватах и проводятся от перехвата к перехвату; это называют салътаторным (скачкообразным) проведением. В этом случае электрический ток течет через внеклеточную жидкость снаружи от миелиновой оболочки, а также через аксоплазму внутри аксона от перехвата к перехвату, последовательно возбуждая один перехват за другим. Таким образом, нервный импульс как будто прыгает по волокну, на основании этого и появился термин «салътаторное проведение».
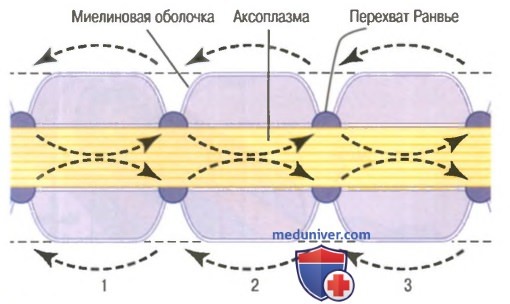
Сальтаторное проведение по миелинизированному аксону. Стрелки указывают направление движения электрического тока от перехвата к перехвату.
Сальтаторное проведение имеет два преимущества. Во-первых, заставляя процесс деполяризации «прыгать» через большие промежутки вдоль аксона, этот механизм повышает скорость проведения в миелинизированных волокнах в 5-50 раз. Во-вторых, сальтаторное проведение сохраняет энергию для аксона, поскольку деполяризуются только перехваты, что позволяет приблизительно в 100 раз снизить потерю ионов по сравнению с возможными потерями в других условиях. В связи с этим снижаются траты энергии, необходимые для восстановления трансмембранной разности концентраций ионов натрия и калия после серии нервных импульсов.
Существует другая особенность сальтаторного проведения в крупных миелинизированных волокнах: отличная изоляция, обеспечиваемая миелиновой оболочкой, и 50-кратное снижение мембранной емкости позволяют осуществлять реполяризацию путем перемещения очень незначительного числа ионов.
Скорость проведения в нервных волокнах. Скорость проведения в нервных волокнах колеблется от 0,25 м/сек в очень тонких немиелинизирован-ных волокнах до 100 м (длина футбольного поля) в 1 сек в очень толстых миелинизированных волокнах.
Видео физиология возбуждения тканей (потенциал покоя, потенциал действия) — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Продолжительность потенциала действия для нервных тканей составляет
Функция нервных клеток в организме заключается в получении информации, передаче ее в другие отделы нервной системы, сопоставление информации от разных источников и, наконец, регуляции деятельности других клеток. Сигналы, поступающие от нервов, вызывают сокращение мышечных клеток. Когда эти два типа клеток «активны» (каждая по-своему), возникает быстрый сдвиг мембранного потенциала в положительном направлении — потенциал действия.
Временной ход потенциала действия
Потенциалы действия можно зарегистрировать в нервных и мышечных клетках с помощью внутриклеточных электродов (рис. 2.1). Типичные примеры потенциалов действия в различных тканях млекопитающих представлены на рис. 2.4. Во всех этих случаях потенциал резко нарастает от отрицательных значений потенциала покоя до положительного пика, составляющего примерно +30 мВ. Затем потенциал с различной скоростью возвращается к уровню покоя; длительность потенциала действия составляет около 1 мс в нервах, 10 мс в скелетной мышце и более 200 мс в миокарде.
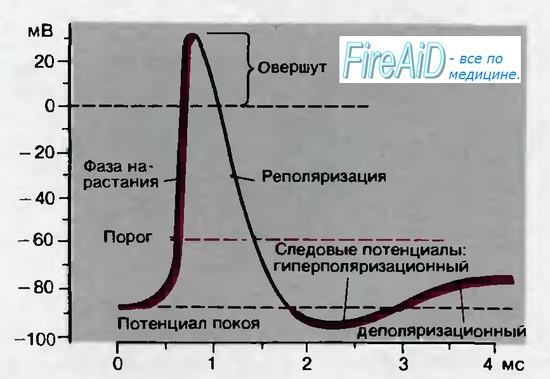
Рис. 2.5. Временной ход потенциала действия в нейроне; показаны последовательные фазы потенциала действия, описанные в тексте
Как показывает рис. 2.5, для потенциала действия характерны несколько фаз. Он начинается очень быстрым сдвигом потенциала в положительном направлении — фазой нарастания, которая продолжается всего лишь 0,2-0,5 мс. Во время фазы нарастания клеточная мембрана теряет свой нормальный заряд («поляризацию»); поэтому фазу нарастания называют также фазой деполяризации. Обычно кривая деполяризации переходит за нулевую линию и мембранный потенциал становится положительным. Эта положительная фаза потенциала действия называется овершутом («перелетом»). Следующая за овершутом фаза, в течение которой восстанавливается исходный потенциал покоя мембраны, называется реполяризацией.
Продолжительность потенциала действия для нервных тканей составляет
В нервных волокнах сигналы передаются с помощью потенциалов действия, которые представляют собой быстрые изменения мембранного потенциала, быстро распространяющиеся вдоль мембраны нервного волокна. Каждый потенциал действия начинается со стремительного сдвига потенциала покоя от нормального отрицательного значения до положительной величины, затем он почти так же быстро возвращается к отрицательному потенциалу. При проведении нервного сигнала потенциал действия движется вдоль нервного волокна вплоть до его окончания.
На рисунке показаны изменения, возникающие на мембране во время потенциала действия, с переносом положительных зарядов внутрь волокна вначале и возвращением положительных зарядов наружу в конце. В нижней части рисунка графически представлены последовательные изменения мембранного потенциала в течение нескольких 1/10000 сек, иллюстрирующие взрывное начало потенциала действия и почти столь же быстрое восстановление.
Читайте также: Хардангер ткань для чего
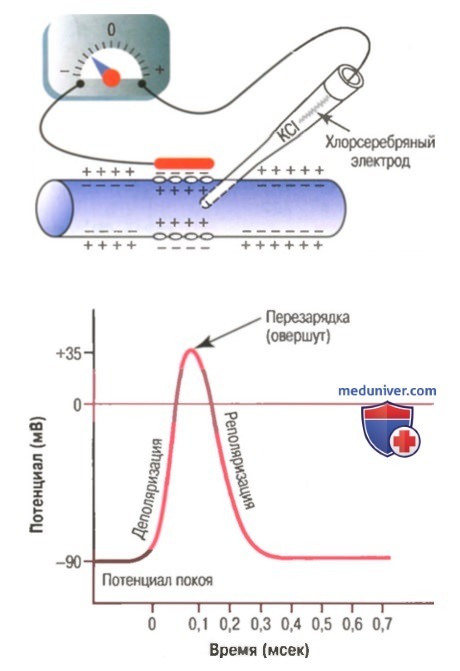
Типичный потенциал действия, зарегистрированный с помощью метода, представленного в верхней части рисунка.
Стадия покоя. Эта стадия представлена мембранным потенциалом покоя, который предшествует потенциалу действия. Мембрана во время этой стадии поляризована в связи с наличием отрицательного мембранного потенциала, равного -90 мВ.
Фаза деполяризации. В это время мембрана внезапно становится высокопроницаемой для ионов натрия, позволяя огромному числу положительно заряженных ионов натрия диффундировать внутрь аксона. Нормальное поляризованное состояние в -90 мВ немедленно нейтрализуется поступающими внутрь положительно заряженными ионами натрия, в результате потенциал стремительно нарастает в положительном направлении. Этот процесс называют деполяризацией, В крупных нервных волокнах значительный избыток входящих внутрь положительных ионов натрия обычно приводит к тому, что мембранный потенциал «проскакивает» за пределы нулевого уровня, становясь слегка положительным. В некоторых более мелких волокнах, как и в большинстве нейронов центральной нервной системы, потенциал достигает нулевого уровня, не «перескакивая» его.
Фаза реполяризации. В течение нескольких долей миллисекунды после резкого повышения проницаемости мембраны для ионов натрия, натриевые каналы начинают закрываться, а калиевые — открываться. В результате быстрая диффузия ионов калия наружу восстанавливает нормальный отрицательный мембранный потенциал покоя. Этот процесс называют реполя-ризацией мембраны.
Для более полного понимания факторов, являющихся причиной деполяризации и реполяризации, необходимо изучить особенности двух других типов транспортных каналов в мембране нервного волокна: электроуправляемых натриевых и калиевых каналов.
Электроупавляемые натриевые и калиевые каналы. Необходимым участником процессов деполяризации и реполяризации во время развития потенциала действия в мембране нервного волокна является электроуправляемый натриевый канал. Электроуправляемый калиевый канал также играет важную роль в увеличении скорости реполяризации мембраны. Оба типа электроуправляемых каналов существуют дополнительно к Na + /K + -насосу и каналам К + /Na + -утечки.
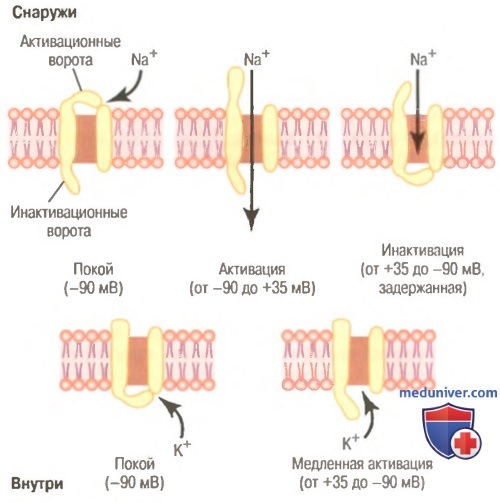
Типичные изменения состояния электроуправляемых натриевых (вверху) и калиевых (внизу) каналов при изменении мембранного потенциала от нормального отрицательного значения до положительных величин.
Показана последовательная активация и инактивация натриевых каналов и задержанная активация калиевых каналов.
Электроуправляемый натриевый канал. В верхней части рисунка показан электроуправляемый натриевый канал в трех различных состояниях. Этот канал имеет двое ворот: одни вблизи наружной части канала, которые называют активационными воротами, другие — у внутренней части канала, которые называют инактивационными воротами. В верхней левой части рисунка изображено состояние этих ворот в покое, когда мембранный потенциал покоя равен -90 мВ. В этих условиях активационные ворота закрыты и препятствуют поступлению ионов натрия внутрь волокна.
Активация натриевого канала. Когда мембранный потенциал покоя смещается в направлении менее отрицательных значений, поднимаясь от -90 мВ в сторону нуля, на определенном уровне (обычно между -70 и -50 мВ) происходит внезапное конформационное изменение актива-ционных ворот, в результате они переходят в полностью открытое состояние. Это состояние называют активированным состоянием канала, при котором ионы натрия могут свободно входить через него внутрь волокна; при этом натриевая проницаемость мембраны возрастает в диапазоне от 500 до 5000 раз.
Инактивация натриевого канала. В верхней правой части рисунке показано третье состояние натриевого канала. Увеличение потенциала, открывающее активационные ворота, закрывает инактивационные ворота. Однако инактивационные ворота закрываются в течение нескольких десятых долей миллисекунды после открытия активационных ворот. Это значит, что конформационное изменение, приводящее к закрытию инактивационных ворот, — процесс более медленный, чем конформационное изменение, открывающее активационные ворота. В результате через несколько десятых долей миллисекунды после открытия натриевого канала инактивационные ворота закрываются, и ионы натрия не могут более проникать внутрь волокна. С этого момента мембранный потенциал начинает возвращаться к уровню покоя, т.е. начинается процесс реполяризации.
Существует другая важная характеристикая процесса инактивации натриевого канала: инактивационные ворота не открываются повторно до тех пор, пока мембранный потенциал не вернется к значению, равному или близкому к уровню исходного потенциала покоя. В связи с этим повторное открытие натриевых каналов обычно невозможно без предварительной реполяризации нервного волокна.
Видео физиология возбуждения тканей (потенциал покоя, потенциал действия) — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в оглавление раздела «Физиология человека.»
Продолжительность потенциала действия для нервных тканей составляет
При образовании потенциалов действия (спайное) в химических синапсах между нейронами происходит высвобождение медиаторов из пузырьков пресинаптической мембраны. Медиатор проходит через синаптическую щель и активирует рецепторы, встроенные в постсинаптическую мембрану нейрона-мишени. Рецепторы, в свою очередь, активируют медиаторозависимые ионные каналы, изменяющие поляризацию нейрона-мишени. При смещении показателей мембранного потенциала от уровня потенциала покоя (-70 мВ) в отрицательную сторону (например, — 80 мВ и более) происходит гиперполяризация мембраны, а при смещении в положительную сторону — деполяризация.
а) Электротонические потенциалы. Первоначальный ответ нейрона-мишени на воздействие возбуждающих импульсов заключается в формировании локальных ступенчатых электротонических потенциалов. Положительные злектротонические потенциалы обычно возникают в мультииолярных нейронах в результате деполяризации, обусловленной активацией медиаторозависимых ионных каналов. При низкочастотном стимулирующем воздействии по дендритам распространяются небольшие волны деполяризации, угасающие в течение 2-3 мс.
При увеличении частоты воздействия происходит ступенчатая временная суммация возбуждения, в результате чего образуется прогрессивно увеличивающаяся волна, которая распространяется на поверхность сомы. Одновременное схождение в области сомы волн возбуждения, распространяющихся по разным дендритам, приводит к пространственной суммации возбуждения. При деполяризации приблизительно на 15 мВ (и значении потенциала -55 мВ) происходит открытие потенциалозависимых ионных каналов в наиболее чувствительной области нейрона — триггерной точке (триггерной области), располагающейся в начальном сегменте аксона. При достижении уровня деполяризации (генераторного потенциала) пороговых значений, т.е. потенциала, способного запустить открытие потенциалозависимых ионных каналов, происходит формирование потенциала действия.
Читайте также: Купальник из ткани лапша
В триггерной зоне чувствительных нейронов черешнях и спинномозговых нервов формируется так называемый рецепторный потенциал. В этой области расположено большое количество ионных каналов, активирующихся под воздействием специфических чувствительных стимулов и вызывающих ступенчатое распространение волны деполяризации.
Определение триггерной области миелинизированных нервных волокон не вызывает затруднений: у мультиполярных нейронов эта область граничит с первым миелиновым сегментом с его проксимальной стороны, а у периферических чувствительных нейронов — с последним миелиновым сегментом с его дистальной стороны.
Тормозные (гиперполяризующие) постсинаптические потенциалы возникают при открытии ионных каналов, генерирующих выходящий ток (например, калиевых каналов). Распространение тормозных потенциалов также является угасающим.
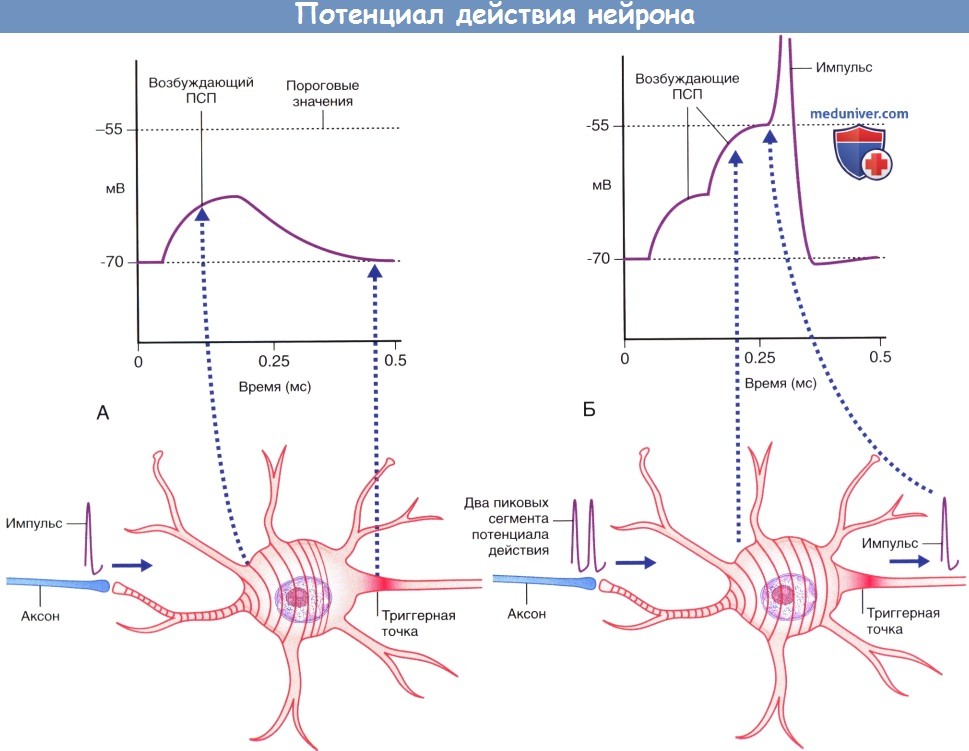 Временная суммация.
Временная суммация.
(А) Аксон чувствительного нейрона (выделен голубым цветом) передает единичный импульс двигательному нейрону,
вызывающий возбуждающему постсинаптический потенциал (ПСП), значения которого не достигают пороговых величин, вследствие чего этот потенциал затухает.
(Б) Аксон чувствительного нейрона передает два импульса.
Происходит временная суммация импульсов с достижением в начальном сегменте аксона пороговых значений возбуждения,
что приводит к формированию потенциала действия, который в дальнейшем распространяется по аксону двигательного нейрона. 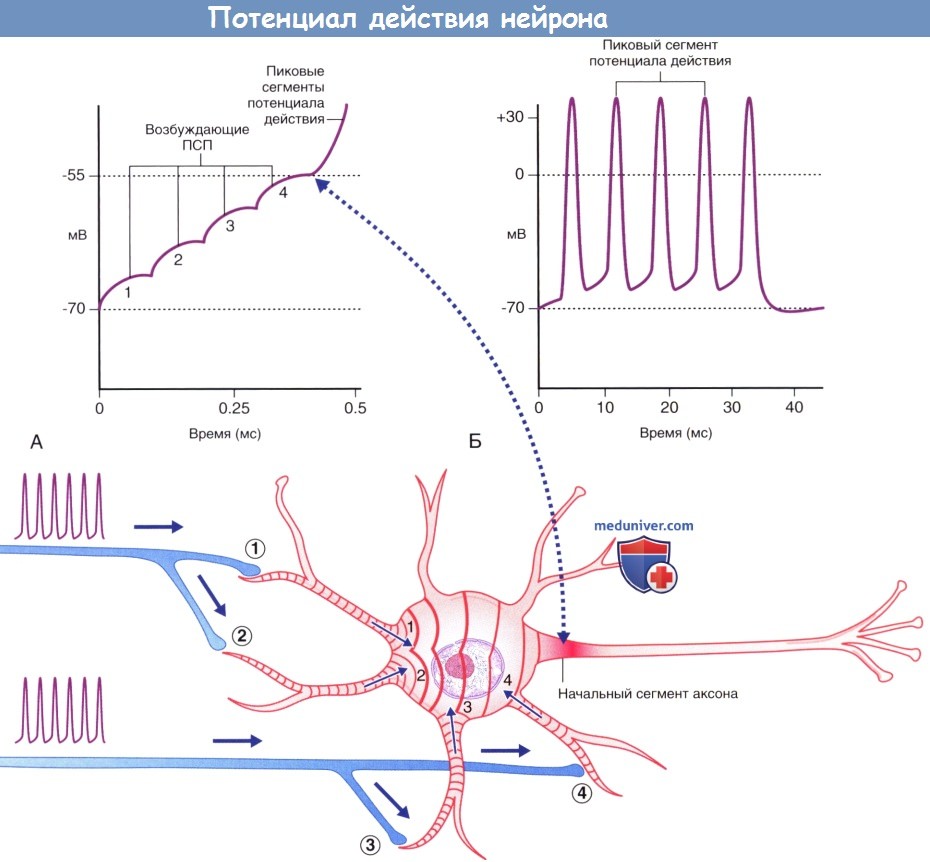 (А) Ступенчатая суммация возбуждающих постсинаптических потенциалов (ПСП), за счет которой происходит формирование пикового потенциала. Пунктирной линией указана область, соответствующая графику А.
(А) Ступенчатая суммация возбуждающих постсинаптических потенциалов (ПСП), за счет которой происходит формирование пикового потенциала. Пунктирной линией указана область, соответствующая графику А.
(Б) Генераторные потенциалы достаточной силы формируют множественные пиковые потенциалы. 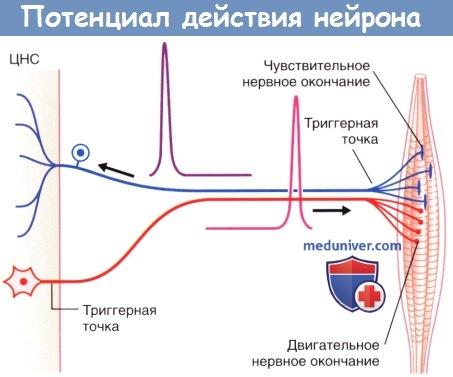 Потенциалы действия двигательных и чувствительных нейронов скелетной мускулатуры. ЦНС — центральная нервная система.
Потенциалы действия двигательных и чувствительных нейронов скелетной мускулатуры. ЦНС — центральная нервная система. 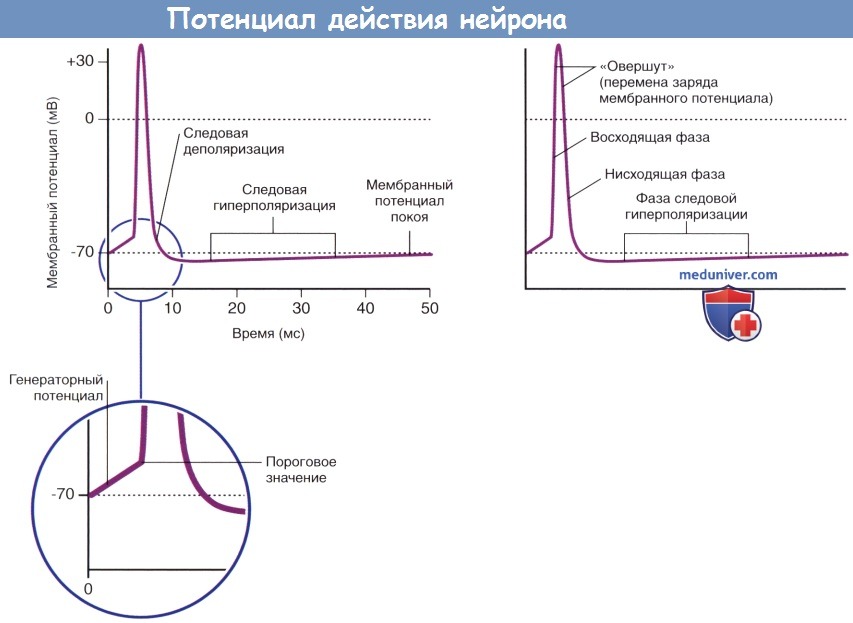 Основные характеристики потенциала действия.
Основные характеристики потенциала действия.
б) Форма потенциала действия. На рисунке ниже представлен единичный потенциал действия. Пиковый сегмент потенциала действия (спайк) начинается при достижении мембранным потенциалом аксонного холмика порогового значения, которое составляет -55 мВ. Восходящая часть спайка (деполяризации мембраны) достигает значения 4-35 мВ, где происходит перемена заряда («овершут»). Во время нисходящей части мембранный потенциал понижается до -75 мВ за счет следовой гиперполяризации, а затем достигает исходных значений.
Деполяризация мембраны до достижения пороговых значений за счет действия медиаторозависимых ионных каналов приводит к открытию потенциалозависимых Na + -каналов. Вход ионов Na + в клетку вызывает деполяризацию и открытие остальных потенциалозависимых Na + -каналов по принципу положительной обратной связи, что приводит к перемене заряда мембранного потенциала 4-35 мВ («овершут») и стремлению к потенциалу Нернста для ионов Na + . В этот момент одновременно начинаются прогрессирующая инактивация натриевых каналов и открытие калиевых каналов. Движение потока ионов Na + в клетку сменяется на выходящий ток ионов К + , что запускает реполяризацию мембраны.
Возникновение фазы следовой гиперполяризации объясняют тем, что после полной инактивации потенциалозависимых натриевых каналов многие калиевые каналы остаются открытыми. Восстановление ионных градиентов обеспечивает натрий-калиевый насос.
В ходе подробного изучения натриевых каналов было обнаружено, что их регуляция осуществляется двумя путями. В состоянии покоя (-70 мВ) активационные ворота, расположенные в средней части натриевых и калиевых каналов, закрыты. Первыми на пороговое раздражение реагируют натриевые каналы: происходят открытие активационных ворот и быстрый выход ионов Na + в клетку по концентрационному и электрическому градиентам. При достижении потенциалом действия пиковых значений (+35 мВ) происходит открытие калиевых каналов, а натриевые каналы закрываются инактивационными воротами, представленными глобулярным белком. При реполяризации и восстановлении мембранного потенциала (-70 мВ) активационные натриевые ворота закрываются, а затем вновь открываются инактивационные натриевые ворота, тем самым приводя натриевые каналы в исходное состояние.
Работа потенциалозависимых калиевых каналов регулирует одноворотный механизм: этими каналами управляют только активационные ворота.
Потенциал действия, возникающий в ответ на деполяризацию, подчиняется принципу «все или ничего», согласно которому при достижении пороговых показателей возбуждения происходит открытие потенциалозависимых натриевых каналов. С этой точки зрения потенциал действия отличается от медиаторозависимых потенциалов, суммирующихся для развития потенциалов действия. Кроме того, отличие потенциала действия от других ступенчатых потенциалов заключается в отсутствии затухания; потенциал действия распространяется в полном объеме от сомы вдоль нервного волокна к нервному окончанию (это расстояние для нейронов нижних конечностей может составлять более метра).
Во время фазы нарастания и в начале фазы реполяризации потенциала действия нейрон находится в абсолютно рефрактерном состоянии и не может воспринимать следующие импульсы в результате инактивации потенциалозависимых натриевых каналов. Вслед за этим периодом наступает интервал относительной рефрактерности, во время которого стимул, превышающий исходный более чем на 15 мВ, способен вызвать формирование повторного потенциала действия. Часто для достижения показателя 35 мВ генераторный потенциал производит от 50 до 100 импульсов в секунду (частота составляет от 50 до 100 Гц).
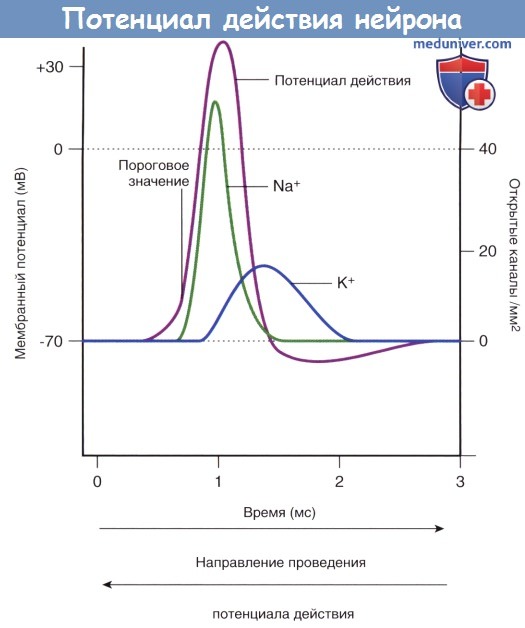 Изменения проницаемости потенциалозависимых натриевых и калиевых каналов, обеспечивающих развитие потенциала действия.
Изменения проницаемости потенциалозависимых натриевых и калиевых каналов, обеспечивающих развитие потенциала действия. 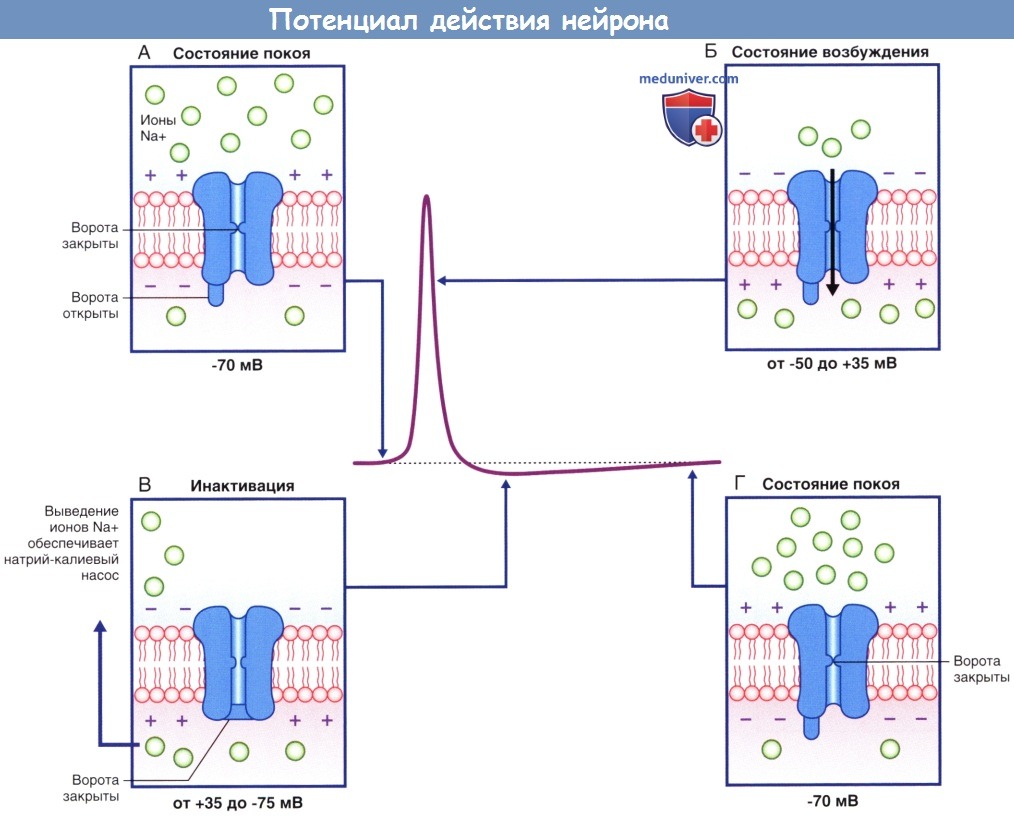 Потенциалозависимые натриевые каналы в разные фазы потенциала действия.
Потенциалозависимые натриевые каналы в разные фазы потенциала действия.
(А) В состоянии покоя, предшествующем возбуждению, активационные ворота находятся в закрытом состоянии, а инактивационные — в открытом.
(Б) При преодолении порога возбуждения происходит открытие активационных ворот, что приводит к полному открытию ионного канала.
(В) Инактивационные ворота закрываются, что приводит к закрытию ионного канала.
(Г) При восстановлении потенциала покоя активационные ворота закрываются, а инактивационные — открываются.
Ионный канал возвращается в исходное состояние. 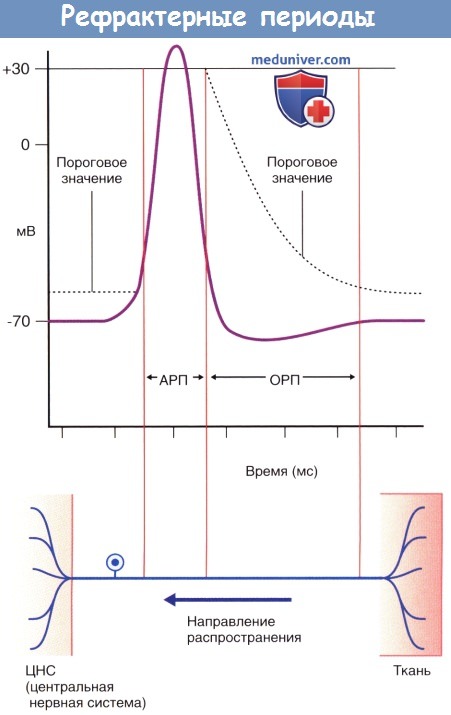 Рефрактерные периоды.
Рефрактерные периоды.
АРП — абсолютный рефрактерный период;
ОРП — относительный рефрактерный период.
в) Передача нервных импульсов. Изменение потенциала триггерной зоны распространяется вдоль аксона в соответствии с электротонической траекторией. Положительные заряды внутренней стороны мембраны распространяются в обоих направлениях через аксоплазму. Положительные заряды внешней стороны мембраны аналогичным образом проходят через внеклеточную жидкость и нейтрализуют отрицательный заряд на внешней стороне мембраны. Прилежащий проксимальный участок мембраны достаточно рефрактерен и способен противостоять деполяризации. Одновременно с этим в прилежащем дистальном сегменте возникает локальный ответ (деполяризация), который достигает пороговых величин и распространяется далее вдоль ствола и ветвей аксона к нервным окончаниям, обеспечивая проведение потенциала действия.
Читайте также: Ткань бежевый жаккард для штор
Проведение импульса по немиелинизированному нервному волокну происходит непрерывно, а по миелинизированному — сальтаторно (скачкообразно). Миелиновая оболочка выполняет функцию изолятора и располагается между перехватами Ранвье, где сконцентрированы натриевые каналы. В связи с этим в каждом перехвате Ранвье последовательно возникают потенциалы действия за счет распространения положительного тока вдоль аксоплазмы миелинизированного участка аксона и его выхода в области следующего перехвата Ранвье. При распространении тока в обратном направлении по внеклеточной жидкости для повторного заряда деполяризованного участка мембраны снятие положительного заряда приводит к деполяризации следующего перехвата Ранвье.
г) Скорость проведения нервных импульсов. Скорость проведения нервного импульса по немиелинизированным нервным волокнам прямо пропорциональна диаметру аксона. В первую очередь это связано с тем, что больший объем аксоплазмы обеспечивает более высокую скорость продольного распространения возбуждения. Кроме того, чем больше диаметр аксона, тем больше площадь поверхности его плазматической мембраны, следовательно, на ней расположено большее количество ионных каналов, а деполяризация и восстановление заряда мембраны происходят быстрее. Диаметр аксонов варьирует от 0,2 до 2 мкм, а скорость проведения импульсов — от 2 до 25 м/с.
Внешний диаметр миелинизированных нервных волокон (с учетом толщины миелинового слоя) варьирует от 2 до 25 мкм. Следует отметить, что чем шире диаметр миелинизированного нервного волокна, тем длиннее миелинизированные участки, расположенные между перехватами Ранвье, в связи с чем потенциалы действия распространяются с большей скоростью, подобно спортсмену, делающему широкие шаги при беге. Скорости проведения нервных импульсов по разным видам периферических нервов описаны отдельной статье на сайте — просим пользоваться формой поиска на главной странице сайта.
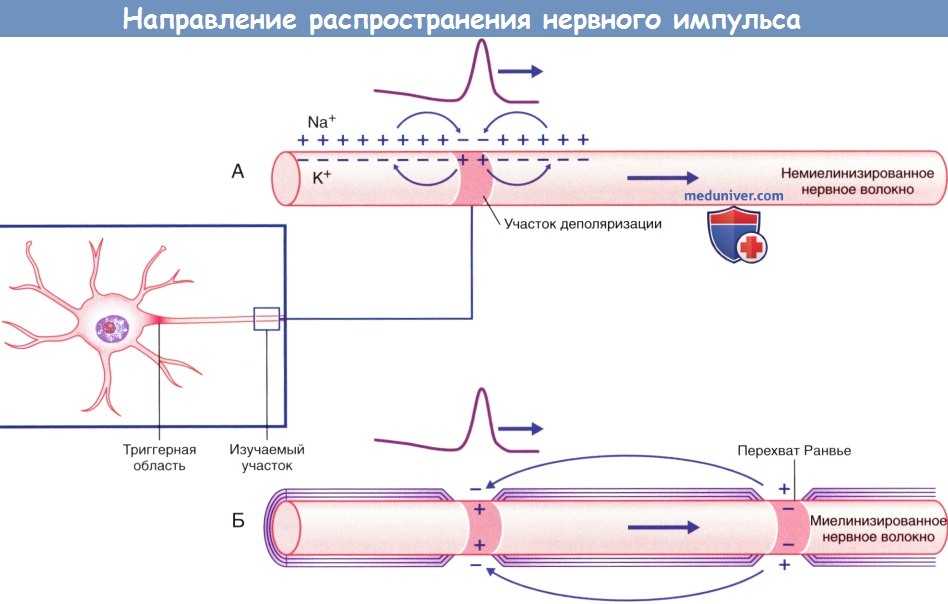
Направление распространения импульса, показанное в виде прохождения положительного заряда.
(А) Непрерывное проведение импульса по немиелинизированному нервному волокну.
(Б) Скачкообразное проведение импульса по миелинизированному нервному волокну.
д) Механизм действия местных анестетиков. При применении местных анестетиков происходит обратимая блокировка проведения нервных импульсов за счет инактивации натриевых каналов, особенно потенциалозависимых, вследствие чего деполяризация мембраны становится невозможной. Молекулы местных анестетиков связывают белок, окружающий ионные каналы. В течение времени действия анестетика происходит связывание белка, окружающего ионные каналы. Для доступа к этим белкам местные анестетики должны быть хорошо растворимы в жирах. При введении анестетика вблизи периферического нерва в первую очередь происходит инактивация немиелинизированных и тонко миелинизированных Аб-волокон. При воздействии на смешанный нерв может возникнуть временный двигательный паралич.
Большинство местных анестетиков — производные амидов (бупивакаин, лидокаин) или сложных эфиров (бензокаин, новокаин). И те, и другие вызывают местное расширение сосудов за счет прямого расслабления гладкой мускулатуры артериол, что приводит к ускорению их выведения. В связи с этим в растворы для местной анестезии часто добавляют адреналин, вызывающий сокращение гладкой мускулатуры и сужение сосудов.
е) Резюме. Электрически заряженные атомы или группы атомов называют ионами. Ионы Na + и К + являются катионами, а ионы Cl + и отрицательно заряженные белки — анионами. Клеточные мембраны представляют собой заряженные конденсаторы с потенциалом покоя величиной -70 мВ.
Неуправляемые натриевые, калиевые и хлорные каналы открыты постоянно, и движение соответствующих ионов через них происходит по градиенту концентрации. Натриевые каналы относительно малочисленны, а калиевые и кальциевые каналы представлены в большом количестве. Ионы К + сконцентрированы в цитозоле за счет притягивающего влияния белков-анионов цитоскелета и отталкивающего действия ионов Na + , расположенных на внешней части плазматической мембраны. Натрий-калиевый насос обеспечивает поддержание мембранного потенциала.
Первоначальная реакция мультиполярного нейрона на возбуждающий стимул заключается в формировании угасающих волн положительного электротонуса. В результате их временной и/или пространственной суммации происходит образование генераторного потенциала в начальном сегменте аксона. При достижении пороговых значений (-55 мВ) потенциалозависимые ионные каналы генерируют потенциал действия, распространяющийся по нервному волокну. Тормозные стимулы вызывают формирование волн отрицательного электротонуса, суммация которых приводит к гиперполяризации мембраны, тем самым отдаляя величины ее потенциала от пороговых значений.
Потенциал действия (спайк) проходит фазу нарастания (деполяризации) от начальных значений до +35 мВ, фазу убывания (реполяризации), в которую возвращается к исходным показателям, и фазу следовой гиперполяризации, в которую понижается до -75 мВ, а затем вновь возвращается к первоначальным значениям. Деполяризация мембраны происходит в результате активации потенциалозависимых натриевых каналов, в ходе которой происходит их стремительное (со скоростью + в клетку и деполяризации мембранного потенциала. При достижении мембранного потенциала +35 мВ происходит закрытие натриевых каналов при помощи инактивационных ворот.
При достижении потенциала действия максимальных значений происходит открытие потенциалозависимых калиевых каналов, и поток ионов Na + в клетку сменяется потоком ионов К + из клетки, что приводит к реполяризации и следовой гиперполяризации мембранного потенциала.
В течение 1 мс после воздействия первоначального импульса триггерная зона начального сегмента аксона характеризуется абсолютной рефрактерностью к восприятию следующих импульсов, а в течение 3 мс—относительной рефрактерностью. Возникновение потенциалов действия подчиняется принципу «все или ничего» и распространяется по всей длине нервного волокна и его ветвям. Проведение нервных импульсов по немиелинизированным аксонам осуществляется непрерывно, а по миелинизированным—сальтаторно (скачкообразно от одного перехвата Ранвье к другому). Сальтаторное проведение характеризуется большей скоростью. Миелинизированные участки длиннее у волокон с большим диаметром, и скорость проведения импульсов по этим волокнам выше.
Редактор: Искандер Милевски. Дата публикации: 11.11.2018
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом