
Культура тканей растений — Plant tissue culture — Wikipedia
Культура тканей растений представляет собой набор методов, используемых для поддержания или выращивания растительных клеток, тканей или органов в стерильных условиях на питательной культуральной среде известного состава. Он широко используется для получения клонов растений методом, известным как микроразмножение. Различные методы культивирования тканей растений могут иметь определенные преимущества по сравнению с традиционными методами размножения, в том числе:
- Производство точных копий растений, которые дают особенно хорошие цветы, плоды или другие желательные качества.
- Для быстрого получения зрелых растений.
- Производство нескольких растений при отсутствии семян или необходимых опылителей для получения семян.
- Регенерация целых растений из генетически модифицированных растительных клеток.
- Производство растений в стерильных контейнерах, что позволяет их перемещать, что значительно снижает вероятность передачи болезней, вредителей и патогенов.
- Получение растений из семян, которые в противном случае имели бы очень низкие шансы на прорастание и рост, т.е. орхидеи и Непентес.
- Для очистки определенных растений от вирусных и других инфекций и для быстрого размножения этих растений в качестве «очищенного стада» для садоводства и сельского хозяйства.
Культура тканей растений основана на том факте, что многие клетки растений обладают способностью регенерировать целое растение (тотипотентность). Одиночные клетки, клетки растений без клеточных стенок (протопласты), кусочки листьев, стеблей или корней часто можно использовать для создания нового растения на питательной среде с учетом необходимых питательных веществ и гормоны растений.
Методы
Подготовка растительной ткани к культуре ткани осуществляется в условиях асептический условия в HEPA фильтрованный воздух, подаваемый шкаф с ламинарным потоком. После этого ткань выращивают в стерильных контейнерах, таких как чашки Петри или колбы в камере выращивания с контролируемой температурой и интенсивностью света. Живые растительные материалы из окружающей среды естественным образом загрязнены на своей поверхности (а иногда и внутри) микроорганизмы, поэтому их поверхности стерилизуют химическими растворами (обычно спиртом и натрий или же гипохлорит кальция) [1] перед подходящими образцами (известными как эксплантаты) принимаются. Затем стерильные эксплантаты обычно помещают на поверхность стерильной твердой культуральной среды, но иногда их помещают непосредственно в стерильную жидкую среду, особенно когда требуются культуры суспензий клеток. Твердые и жидкие среды обычно состоят из неорганический соли плюс несколько органических питательных веществ, витаминов и гормонов растений. Твердый средства массовой информации готовятся из жидких сред с добавлением гелеобразующего агента, обычно очищенного агара.

Состав среды, особенно растительные гормоны и источник азота (нитрат по сравнению с солями аммония или аминокислотами), оказывают сильное влияние на морфологию тканей, которые растут из исходного эксплантата. Например, превышение ауксин часто приводит к разрастанию корней, в то время как избыток цитокинин может давать всходы. Баланс ауксина и цитокинина часто вызывает неорганизованный рост клеток или мозоль, но морфология нароста будет зависеть от вида растения, а также от состава среды. По мере роста культур кусочки обычно отрезают и пересеивают на новую среду, чтобы обеспечить рост или изменение морфологии культуры. Навыки и опыт специалиста по культуре тканей важны при решении, какие кусочки культивировать, а какие выбросить.
По мере прорастания побегов из культуры их можно срезать и укоренять с помощью ауксина для получения проростков, которые, когда они созреют, могут быть перенесены в почву для дальнейшего роста в теплице как нормальные растения. [2]
Пути регенерации

Специфические различия в регенерационном потенциале разных органов и эксплантов имеют разные объяснения. Существенные факторы включают различия в стадии клеток в клеточный цикл, наличие или способность транспортировать эндогенные регуляторы роста и метаболические возможности клеток. Чаще всего используются тканевые эксплантаты. меристематический концы растений, такие как кончик стебля, кончик пазушных почек и кончик корня. Эти ткани обладают высокой скоростью деления клеток и либо концентрируют, либо производят необходимые регулирующие рост вещества, включая ауксины и цитокинины.
Эффективность регенерации выстрела в культура ткани обычно количественная характеристика который часто варьируется между видами растений и внутри одного вида растений среди подвидов, разновидностей, сорта, или же экотипы. Следовательно, регенерация тканевой культуры может стать сложной, особенно когда необходимо разработать множество процедур регенерации для разных генотипы внутри одного вида.
Три общих пути регенерации культуры тканей растений — это размножение из уже существующих меристем (культура побегов или узловая культура), органогенез и незиготный эмбриогенез.
Размножение побегов или узловых сегментов обычно выполняется в четыре этапа для массового производства проростков через in vitro Вегетативное размножение, но органогенез — это распространенный метод микроразмножения, который включает регенерацию тканей придаточных органов или пазушных зачатков непосредственно или косвенно из эксплантатов. Незиготный эмбриогенез — заслуживающий внимания путь развития, который очень сравним с таковым у зиготических эмбрионов, и это важный путь для производства сомаклональных вариантов, развития искусственных семян и синтеза метаболитов. Из-за одноклеточного происхождения незиготных зародышей они предпочтительны в нескольких системах регенерации для микроразмножения, манипуляции с плоидностью, переноса генов и производства синтетических семян. Тем не менее, регенерация тканей с помощью органогенеза также оказалось полезным для изучения регуляторных механизмов развития растений.
Выбор эксплантата
Ткань, полученная из культивируемого растения, называется эксплантатом.
Эксплантаты могут быть взяты из многих различных частей растения, включая части побегов, листьев, стеблей, цветов, корней, отдельных частей. недифференцированные клетки и из многих типов зрелых клеток при условии, что они все еще содержат живую цитоплазму и ядра и способны к дедифференцировке и возобновлению деления клеток. Это дало начало концепции тотипотентности растительных клеток. [3] [4] Однако это верно не для всех клеток или для всех растений. [5] У многих видов эксплантаты различных органов различаются по скорости роста и регенерации, а некоторые вообще не растут. Выбор материала эксплантата также определяет, являются ли проростки, выращенные в культуре ткани, гаплоидный или же диплоид. Кроме того, риск микробного заражения увеличивается при использовании неподходящих эксплантов.
Первый метод, включающий меристемы и индукцию множественных побегов, является предпочтительным методом для индустрии микроразмножения, поскольку риски сомаклональной изменчивости (генетическая изменчивость, индуцированная в культуре ткани) минимальны по сравнению с двумя другими методами. Соматический эмбриогенез — это метод, у которого есть потенциал в несколько раз выше по скорости размножения, и его можно использовать в системах жидких культур, таких как биореакторы.
Некоторые эксплантаты, например кончик корня, трудно изолировать и заражены почвенной микрофлорой, которая становится проблематичной в процессе культивирования тканей. Определенная микрофлора почвы может образовывать тесные ассоциации с корневые системы, или даже расти внутри корня. Частицы почвы, прикрепленные к корням, трудно удалить, не повредив корни, что может привести к атаке микробов. Эти связанные микрофлора обычно разрастается в среде для культивирования тканей до того, как произойдет значительный рост растительной ткани.
Некоторые культивируемые ткани медленно растут. Для них будет два варианта: (i) Оптимизация питательной среды; (ii) Выращивание высокочувствительных тканей или разновидностей. [6] Некроз может испортить культивируемые ткани. Как правило, сорта растений различаются по восприимчивости к некрозу культур тканей. Таким образом, с этим можно справиться путем культивирования высокочувствительных сортов (или тканей). [6]
Воздушные (надземные) эксплантаты также богаты нежелательной микрофлорой. Однако их легче удалить из эксплантата осторожным промыванием, а оставшуюся часть обычно можно убить стерилизацией поверхности. Большая часть поверхностной микрофлоры не образует прочных ассоциаций с ткань растения. Такие ассоциации обычно можно обнаружить при визуальном осмотре в виде мозаики, обесцвечивания или локализации. некроз на поверхности эксплантата.
Альтернативой для получения незагрязненных эксплантатов является взятие эксплантов из сеянцев, выращенных в асептических условиях из семян, стерилизованных с поверхности. Твердая поверхность семян менее проницаема для агрессивных поверхностных стерилизующих агентов, таких как гипохлорит, поэтому приемлемые условия стерилизации, используемые для семян, могут быть намного более жесткими, чем для вегетативных тканей.
Культурные растения клоны. Если исходное материнское растение, используемое для получения первых эксплантатов, восприимчиво к патогену или условиям окружающей среды, вся культура будет подвержена той же проблеме. И наоборот, любые положительные черты также останутся в рамках этой линии.
Приложения
Культура тканей растений широко используется в растениеводстве, лесоводстве и садоводстве. Приложения включают:
- Коммерческое производство растений, используемых в качестве горшечных растений, ландшафта и цветоводства, при котором используются меристемы и культура побегов для получения большого количества идентичных особей.
- К сохранить редкие или исчезающие виды растений. [7]
- А селекционер может использовать тканевую культуру для скрининга клеток, а не растений на наличие полезных признаков, например гербицид сопротивление / терпимость.
- Масштабный рост растительных клеток в жидкой культуре в биореакторы для производства ценных соединений, таких как вторичные метаболиты растительного происхождения и рекомбинантные белки используется как биофармацевтические препараты. [8]
- Скрещивать отдаленно родственные виды слияние протопластов и возрождение романа гибридный.
- Быстро изучить молекулярные основы физиологических, биохимических и репродуктивных механизмов растений, например, отбор растений, устойчивых к стрессу, in vitro. [9]
- Для перекрестного опыления отдаленно родственных видов и затем культивирования тканей полученного эмбриона, который в противном случае обычно погиб бы (Спасение эмбрионов).
- Для удвоения хромосом и индукции полиплоидия, [10] например удвоенные гаплоиды, тетраплоиды, и другие формы полиплоиды. Обычно это достигается применением антимитотические агенты Такие как колхицин или же Оризалин.
- В качестве ткани для трансформации с последующим краткосрочным тестированием генетических конструкций или регенерацией трансгенный растения.
- Некоторые методы, такие как культивирование верхушек меристемы, можно использовать для получения чистого растительного материала из вирусов, таких как сахарный тростник, [11] картофель и многие виды мягких фруктов.
- Может быть получено производство идентичных стерильных гибридных видов.
- Крупномасштабное производство искусственных семян посредством соматического эмбриогенеза [12]
- Синтетические семена — соматический зародыш инкапсулируется искусственным эндоспермом и искусственной оболочкой семян.
Лаборатории
Хотя некоторые производители и питомники имеют свои собственные лаборатории для размножения растений методом тканевой культуры, ряд независимых лабораторий предоставляют услуги по размножению на заказ. В разделе «Обмен информацией о культурах тканей растений» перечислены многие коммерческие лаборатории по культуре тканей. Поскольку культивирование тканей растений — очень трудоемкий процесс, это будет важным фактором при определении того, какие растения будут коммерчески жизнеспособными для размножения в лаборатории.
Метод культуры тканей. Источник получения растительного сырья
» data-shape=»round» data-use-links data-color-scheme=»normal» data-direction=»horizontal» data-services=»messenger,vkontakte,facebook,odnoklassniki,telegram,twitter,viber,whatsapp,moimir,lj,blogger»>
Метод культуры тканей. КУЛЬТУРА ТКАНЕЙ И КЛЕТОК ЛЕКАРСТВЕННЫХ РАСТЕНИЙ — НОВЫЙ ИСТОЧНИК ПОЛУЧЕНИЯ ЛЕКАРСТВЕННОГО РАСТИТЕЛЬНОГО СЫРЬЯ
В решении задач расширения источников получения ЛРС, повышения стабильности и импортозамещения сырьевой базы перспективным направлением представляется метод биотехнологии, основанный на выращивании клеток и тканей ЛР на искусственных питательных средах. Биотехнологические способы получения массы клеток ЛР возникли на основе развития метода культуры тканей. Под «культурой тканей растений» принято понимать выращивание in vitro (в стерильных искусственных условиях) изолированных клеток, тканей, органов и их частей.
Читайте также: Что можно сшить из тентовой ткани
Метод культуры тканей возник как экспериментальная биологическая модель, позволяющая изучать физиологические, биохимические и другие процессы на уровне автономных клеток, освобожденных от регулирующего влияния целого растительного организма.
История развития метода культуры ткани начинается в XX в. с опытов немецкого ученого Г. Габерландта (1902), впервые высказавшего идею о возможности выращивания клеток, изолированных из организма. Фундаментальные исследования Ф. Уайта (1931, США) и Р. Готре (1932, Франция) позволили определить условия для воспроизведения деления и роста клеток в культуре, и метод культуры тканей приобрел современные черты. В последующие годы были разработаны технические основы метода: отработана методика вычленения тканей и клеток из растений, получения каллусов, сохранения стерильности, усовершенствованы составы питательных сред. В результате этого стало возможным использовать метод культуры тканей для длительного выращивания недифференцированных растительных клеточных масс — каллусов, затем был разработан метод выращивания растительных клеток в суспензионной культуре и получения биомассы от единичных клеток, что позволило выделять однородный в генетическом и физиологическом отношении материал.
Первыми ЛР, которые исследовали в культуре ткани, были барвинок розовый и белена черная, причем Й. Телле и Р. Готре (1947) опытами доказали способность культуры ткани белены к синтезу алкалоидов. Эти вещества накапливались в калуссной массе, получившейся в результате разрастания тканей растения, а также обнаруживались в среде культивирования. Таким образом, культивируемые растительные ткани могли использоваться для производства ЛС. Позже появились сообщения о культуре тканей других ЛР, синтезирующих уникальные продукты.
В 50-х гг. XX в. стали выходить публикации о выращивании растительных тканей в виде суспензионной культуры в жидкой питательной среде, что свидетельствовало о постепенном переходе к получению больших количеств биомассы в специальной аппаратуре — хемостатах, ферментерах, турбидостатах. В это же время наметились области применения клеточных культур в фармацевтической промышленности.
Первоначально метод культуры тканей разрабатывался как чисто теоретическое направление, а с середины 1960-х гг. он вошел в арсенал особой научно-производственной деятельности, известный под названием «биотехнология». Технологии, основанные на методе культуры тканей, помогают создавать новые формы и сорта сельскохозяйственных и ЛР и получать промышленным путем продукты растительного происхождения.
В СССР системные исследования в этой области начались в 1957 г. в Институте физиологии растений АН СССР ученым Р. Г. Бутенко. В 1965 г. по инициативе профессора И. В. Грушвицкого при кафедре фармакогнозии Ленинградского химико-фармацевтического института была создана лаборатория по изучению ЛР в культуре in vitro. Основными объектами изучения как возможных продуцентов препаратов для лечения сердечно-сосудистых заболеваний явились культуры тканей тропических видов раувольфии и растения сем. Аралиевые, известные своими тонизирующими и адаптогенными свойствами. Позже подобные лаборатории были организованы в ВИЛАРе (Москва), Томском медицинском институте, Харьковском химико-фармацевтическом институте и ряде других учреждений.
До 1970-х гг. спектр соединений, которые образовывались культурами тканей в количествах, характерных для целого растения, был ограничен. Это никотин, в больших количествах (0,7 %) синтезируемый клетками табака, диосгенин в культуре диоскореи (1,6 %), виснагин, содержание которого в каллусе амми зубной было в 20 раз больше, чем в растении.
Экспериментальные данные, накопившиеся к данному периоду, указывали, что биосинтез многих соединений в недифференцированных тканях сильно репрессирован, а появление вторичных продуктов во многих случаях было связано с регенерацией корней, побегов и других морфологических структур, т. е. с процессом дифференцировки тканей.
С начала 1970-х гг. список фармакологически ценных вторичных продуктов биосинтеза, обнаруженных в культурах тканей, значительно расширился. Способность некоторых культур к образованию соединений, не обнаруженных в исходных растениях, позволила рассматривать их как продуценты принципиально иных, нетрадиционных БАВ (убихинон-10 в культуре ткани табака, антраценгликозиды в культурах ткани следующих видов: хинное дерево и наперстянка, алкалоиды перакин, вомиленин и другие в культуре ткани раувольфии змеиной). Эти факты указывают на возможность направленного синтеза природных соединений в культуре ткани путем введения в состав питательной среды простых, доступных соединений для их биотрансформации ферментной системой культур тканей в ценные ФАВ. В настоящее время подготовлена к промышленному использованию технология биотрансформации карденолида дигитоксина в дорогостоящий дефицитный дигоксин.
В 1983 г. японская фирма Mitsui Petrochemical Industries опубликовала технологию получения шиконина с помощью культуры клеток воробейника красного (Lithospermum erythrorhizonSieb. et Zucc.), что явилось началом эры биотехнологии, когда биотехнологическое использование культур клеток и тканей в качестве сырья в промышленных масштабах стало реальностью.
В России широкое производство продуктов культуры ткани растений началось с выпуска экстракта культивируемой биомассы женьшеня. Экстракт биомассы женьшеня, или препарат Биоженьшень, стали использовать в качестве БАД к кремам, лосьонам, а в пищевой промышленности — для приготовления тонизирующих напитков. Фармакологический комитет при МЗ РФ разрешил применение настойки из биоженьшеня в качестве аналога по действию корня женьшеня. В г. Харькове (Украина) из биомассы культуры ткани раувольфии змеиной (Rauwolfia serpentina Benth.) было налажено производство ценного антиаритмического ЛС Аймалин.
Культивирование растительных клеток и тканей на искусственной питательной среде в биореакторах помогает решить многие экономические, экологические и технологические задачи, а также преодолеть ряд проблем:
- -свести к минимуму влияние географических, климатических, сезонных, эдафических и прочих условий;
- -добиться стандартности накапливаемых БАВ;
- -регулировать процесс биосинтеза БАВ с использованием разных технологических режимов;
- -выращивать культуры на малых площадях и использовать базу и технологии для синтеза практически всех классов ФАВ в дальнейшем;
- -научиться получать ФАВ, свойственные интактному растению (никотин, кодеин, хинин, диосгенин), и синтезировать новые БАВ;
- -изучить возможность использования культуры растительных клеток для биотрансформации БАВ в конечные ЛС;
- -получить возможность промышленного производства биомассы экзотических растений, малодоступных для нашей страны, например таких, как раувольфия, диоскорея, унгерия и др.;
- -добиться экономической рентабельности биотехнологического производства ФАВ и сокращения посевных площадей под ЛР.
В 1980-хгг. на базе метода культуры ткани возникли новые направления биотехнологии, важнейшим из которых была клеточная инженерия. Изучалось поведение отдельных изолированных клеток в культуре, воздействие на клетки мутагенных факторов и условий внешней среды для получения новых форм растений, получение гибридных растений с помощью протопластов (частей клеток, лишенных оболочки). Способность клеток в культуре тканей при изменении условий культивирования давать начало целому растению привела к созданию промышленных клеточных технологий микроклонального размножения растений, позволяющих в короткие сроки (2—3 мес., а не несколько лет, затрачиваемых при использовании обычных методов) размножать ценные генотипы.
Наряду с культурами клеток и тканей растений развиваются способы культивирования органов растений in vitro (например, культивирование волосатых корней, измененных с помощью бактерий рода Agrobacterium,в качестве альтернативного источника продуктов жизнедеятельности растений, где по условиям роста и скопления клеток возникают субпопуляции с повышенной дифференцировкой, — самые продуктивные клетки по БАВ).
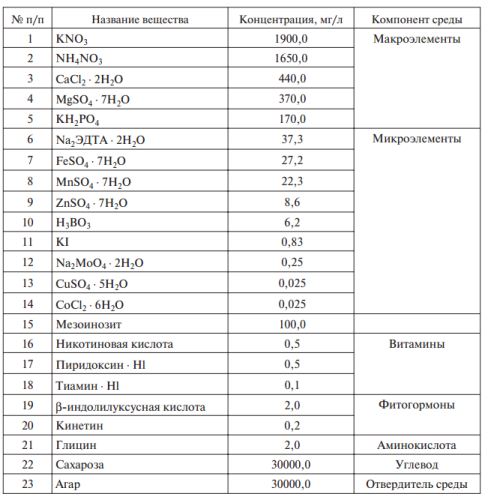
Каллусная и клеточная суспензионная культуры растений Методика получения культуры ткани хорошо разработана и обычно не вызывает вопросов. Чтобы получить культуру ткани, из любой части растения вычленяют эксплант (кусочек ткани размером 0,5—1,0 см, из образовавшегося каллуса для пересева размером 2—4 мм) и помещают на питательную среду (табл. 1, рис. 2).
Таблица 1. Состав среды, в мг/л (Т. Мурасиге и Ф. Скуг, 1962)
Ткани, культивируемые in vitro, перед помещением на питательную среду должны быть стерильными. Стерилизуются исходные кусочки ткани растений, питательная среда; асептически в специальных боксах стерильным инструментом проводятся манипуляции с выращиванием объектов. Чашки Петри, пробирки, пипетки, колбы и иные сосуды, в которых культивируются ткани и клетки, закрываются так, чтобы предотвратить инфицирование в течение продолжительного времени.
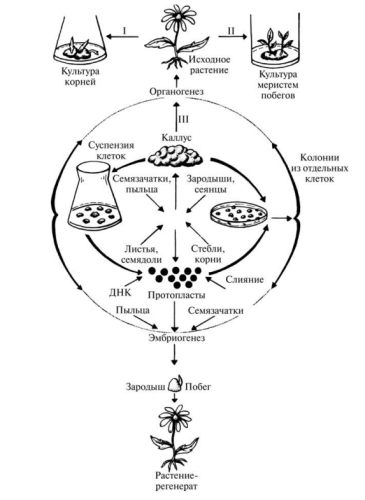
Рис. 2. Схема получения каллусной и суспензионной культур растений, а также из них — растений-регенератов
Асептика является обязательной и необходимой для культивирования как отдельных клеток, так и фрагментов ткани или органа растения (экспланта). Эпифитная микрофлора на тканях растений позже может обнаружиться в культуре ткани. Внутреннее инфицирование растительной ткани чаще всего встречается у тропических и субтропических ЛР. Поэтому кроме поверхностной стерилизации с использованием дезинфицирующих веществ применяют антибиотики, убивающие микробную флору внутри ткани, однако нужно подбирать антибиотик направленного действия.
Наиболее популярна среда, разработанная в 1962 г. Т. Мурасиге и Ф. Скугом. На данной среде можно инициировать и поддерживать рост большого числа культур растительных тканей. Основу питательных сред составляют минеральные соли — макроэлементы (азот в нитратной и аммонийной форме, соли К, Mg, фосфаты и др.) и микроэлементы (Сu, Со, Мо, В, I и др.), дополняемые углеводами, витаминами, стимуляторами роста.
Каллусы легко образуются на эксплантах, взятых из различных органов и частей растений: отрезков стебля, листа, корня, проростков семян, фрагментов паренхимы, тканей клубня, органов цветка, плодов, зародышей и т. д. В природе каллусообразование — естественная реакция на повреждение растений. В культуре изолированных тканей при помещении экспланта на питательную среду его клетки дедифференцируются и переходят к делению, формируя в течение нескольких дней однородную бесформенную массу серо-белого или желтого цвета — каллус. Формирование каллуса длится обычно 1—2 мес. Образовавшийся каллус в асептических условиях разделяют и переносят на свежую питательную среду. Пересаженные ткани растут в контролируемых условиях при температуре 24—28-С. Периодичность субкультивирования тканей зависит от скорости роста биомассы.
Каллусная клетка развивается аналогично другим клеткам, проходя соответственно такие циклы, как деление, растяжение, дифференцировка, старение и отмирание. Кривая роста каллусной ткани имеет S-образный характер и включает пять фаз разной длительности у разных растений:
- -латентная (лаг-фаза — клетки адаптируются и готовятся к делению);
- -линейная (рост каллусной ткани идет с постоянной скоростью);
- -экспоненциальная (время максимальной митотической активности — рост клетки ускорен, масса каллуса увеличивается);
- -стационарная (интенсивность деления резко снижается);
- -отмирания.
Стабильность синтеза вторичных метаболитов как целевого продукта зависит главным образом от стадии культивирования и дифференциации клеток. Однако на вопрос, как связан синтез вторичных метаболитов с ростовыми процессами, ответа пока нет. У большого числа культур вторичные метаболиты синтезируются и накапливаются в значительных количествах либо во время экспоненциальной фазы, когда ростовые процессы особенно активны, либо в период стационарной фазы роста культуры клеток, когда прирост клеточной массы прекращается. Тем не менее есть культуры (например, барвинок розовый —Catharanthus roseus(L.) G. Donf.), у которых синтез вторичных метаболитов происходит в течение всего периода роста.
Синтез вторичных соединений может коррелировать с процессом дифференциации в культуре клеток. Например, в суспензионной культуре мака снотворного (Papaver somniferumL.) синтез алкалоидов начинается после того, как в ней дифференцируется большое количество специализированных клеток млечников, предназначенных для депонирования метаболитов. Синтез вторичных метаболитов в культивируемых клетках связан в основном с пластидами и эндоплазматическим ретикулумом. В клетках, не способных к транспорту метаболитов, продукты вторичного синтеза, как правило, накапливаются в вакуолях и свободном пространстве.
Читайте также: Как сшить фартук из джинсовой ткани
Отметим, что клетки каллусной культуры обычно не транспортируют синтезируемые метаболиты в питательную среду или другие клетки, хотя некоторые культуры являются исключением, в частности культура клеток мака, которые депонируют алкалоиды в млечники. Культивирование тканей растений можно осуществлять на агаризованных питательных средах, имеющих плотную консистенцию, или в жидкой среде. В первом случае ткани образуют скопление недифференцированных клеток, называемых каллусом или биомассой, во втором — клетки при размножении образуют суспензии. Из сравнения каллусных и суспензионных культур следует, что выход продуктов вторичного метаболизма выше в каллусных культурах, но при этом управление процессом культивирования легче осуществлять при работе с суспензионными культурами. Использование технологий получения каллусных культур из ЛРС дает такие преимущества, как надежность и стабильность биомассы и выхода продуктов вторичного метаболизма, а также возможность использования каллусной системы для иммобилизации с последующей биотрансформацией.
Развитие суспензионного метода выращивания (в жидкой питательной среде) позволило превратить культуры тканей растений в удобную модель для исследований. Разрабатываются способы культивирования, сочетающие применение жидкой питательной среды и твердого субстрата, поддерживающего тканевую массу на поверхности, — так называемые иммобилизованные клеточные культуры. В качестве подложки могут использоваться гели из агарозы, альгината, нейлона, полиуретана, полиакриламида, шарики из стекла и др. Иммобилизованные каллусные клетки прекращают рост, но продолжают синтез метаболитов, выделяя их в среду. Основные преимущества иммобилизации — выделение клетками метаболитов в питательную среду, из которой их легко извлечь. Кроме того, иммобилизованные клеточные культуры растений часто используют для биотрансформации.
Довольно часто синтез метаболитов в суспензионной культуре останавливается на промежуточных этапах, не давая необходимого конечного продукта. В этом случае для получения конечного продукта необходима биотрансформация этих метаболитов с помощью культур других растений (или даже клеток бактерий) в целях повышения биологической активности конкретной химической структуры. Так, растения наперстянки шерстистой (Digitalis lanataEchrh.) в большом количестве синтезируют дигитоксин, а не дигоксин. Для соответствующей биотрансформации с успехом используют недифференцированную суспензионную культуру наперстянки, которая с помощью ферментов осуществляет необходимое превращение БАВ. Другой пример: клетки суспензионной культуры из корня женьшеня (Panax ginsengC. A. Mey.) способны биотрансформировать (гликозилировать) фенольные соединения — продукты жизнедеятельности клеток его каллусной культуры.
Таким образом, культура ткани представляет собой периодически пересаживаемую бесформенную массу клеток, способную к неограниченному росту и синтезу специфических продуктов (см. рис. 2). Внешне каллусная ткань совершенно не похожа на растение, от которого она была получена, но ее клетки, как и все клетки растения, несут генетическую информацию, свойственную данному виду (тотипотентность). Процессы, происходящие в культивируемых каллусах, в принципе не отличаются от процессов, идущих в тканях целого растения. Сохраняющаяся в каллусных клетках способность к синтезу специфических вторичных метаболитов — алкалоидов, стероидов (карденолидов, сапонинов), терпеноидов (эфирных масел) и др. — определяет практическую ценность культур растительных тканей для создания технологий промышленного выращивания биомассы клеток в качестве принципиально нового вида ЛРС. В настоящее время технологии, основанные на культивировании тканей высших растений в целях получения редких и дорогостоящих веществ, включены в биотехнологические программы во многих странах.
В исследованиях по культуре тканей ЛР можно выделить три главных направления: получение недифференцированной каллусной массы, создание исходного генетического разнообразия форм растений, а также клеточную селекцию и клональное микроразмножение растений.
Факторы, влияющие на продуктивность культур тканей
Способность культур тканей к накоплению вторичных продуктов обмена является уже установленным фактом. Однако, как правило, клеточные культуры характеризуются низким содержанием требуемых веществ. Чтобы служить ЛРС, в котором содержание вторичных метаболитов достаточно велико, культуры тканей растений необходимо изучить, а затем отобрать высокопроизводительные штаммы.
Тем не менее для многих культур попытки ученых определить условия накопления БАВ, характерных для родительских растений, остаются безрезультатными. В частности, в каллусной культуре не удается получить накопление эфирных масел, которые в естественных условиях синтезируются в особых железках на эпидермисе. Нередко культуры тканей продуцируют вещества иной природы, чем интактные растения: так, коробочки мака снотворного — источники получения морфина, но культура ткани этого растения под влиянием элиситоров образует сангвинарин. Клетки хинного дерева (Cinchona ledgerianaMoens ex Trimen) в культуре накапливают не алкалоиды, а антрахиноны. На выход вторичных продуктов в культуре ткани влияют следующие факторы: происхождение ткани, условия культивирования, клеточная дифференциация in vitro.
Происхождение ткани. Обычно для введения в культуру ткани проводят поиск наиболее продуктивных растений с целью, чтобы эта способность была перенесена и в культуру. Указанный вопрос обсуждается до сих пор, так как экспериментальные данные довольно противоречивы. Например, культуры тканей барвинка (катаранта) розового, полученные из высокоалкалоидных растений, имели тенденцию к синтезу большего количества алкалоидов, чем культуры, полученные от малопродуктивных растений. Наибольшее содержание тропановых алкалоидов (0,02 %) отмечено в культуре ткани белены черной, хотя растение не дает такой высокой продуктивности алкалоидов. В то же время клеточные культуры растения дюбуасия наркотическая (Duboisia myoporoides R. Br.), наиболее богатого тропановыми алкалоидами (5 %), не накапливали значительного количества алкалоидов.
Условия культивирования.
Питание. Важнейшим фактором создания эффективной биотехнологической системы является разработка питательной среды, которая обеспечивает потребности продуцента в химических компонентах, требуемых для оптимального синтеза целевого продукта. Часто при введении в культуру тканей нового вида растений исследователь испытывает большое количество сред, рекомендованных для других видов растений. Этот процесс занимает длительное время и часто не приносит ожидаемого успеха. В последнее время при оптимизации питательных сред используются методы математического планирования биологического эксперимента, которые обладают большой эффективностью и позволяют в короткие сроки подобрать питательные среды, способствующие высокой продуктивности культуры ткани.
Источник углерода (обычно сахароза) — важнейший компонент среды и должен усваиваться клетками растений, переходящими на гетеротрофный тип питания. Для включения автотрофного питания необходимы гормональные и иные изменения состава среды и фотоэкспозиция культур. Однако у многих видов получение фотосинтезирующих культур, эмбриоидов и регенерация культур в растение сопровождается определенными трудностями.
В среде, где все питательные вещества в избытке, увеличение концентрации сахарозы, как правило, приводит к увеличению биомассы. В некоторых случаях увеличение сахарозы может оказать положительный эффект и на выход действующих веществ. Так, в культуре ткани катаранта розового при увеличении концентрации сахарозы в 4 раза концентрации серпентина и антоцианов увеличились в 2,6 раза. Концентрация алкалоидов в клетках растений эшшольция (EschscholtsiaCham.) увеличивалась в 7 раз при увеличении концентрации сахарозы в 4 раза. Но содержание фенольных соединений, продуцируемых этими клетками, не изменилось, что указывает на независимую регуляцию различных биосинтетических путей.
Для некоторых культур разработаны способы двухэтапного выращивания, когда ткани после накопления достаточной биомассы переносят в продукционные среды, способствующие максимальному синтезу БАВ. Примерами эффективных продукционных сред являются среда А. Г. Волосовича и других(1982), а также жидкая питательная среда, содержащая 8 % сахарозы, без фитогормонов с уменьшенным содержанием фосфора, обеспечивающая в 4—8 раз более высокую продукцию алкалоидов клетками суспензионной культуры эшшольция, чем целого растения.
Стрессовые факторы. Количество образованных вторичных продуктов в культуре ткани может резко возрастать под влиянием некоторых стрессовых факторов (воздействие продуктов жизнедеятельности микроорганизмов, осмотического шока, токсических ионов тяжелых металлов и т. д.). Вторичные продукты растений часто являются фитоалексинами, и их синтез в растительной клетке происходит в ответ на действие продуктов жизнедеятельности микроорганизмов для защиты от фитопатогенов. При добавлении к культуре ткани элиситоров (компоненты стенок грибного мицелия или какие-либо другие продукты жизнедеятельности микроорганизмов) интенсивность синтеза клетками растений некоторых фармакологически ценных веществ возрастает. Поэтому Институт биотехнологии (г. Саскачеван, Канада) в целях уменьшения себестоимости конечного продукта предложил промышленный способ получения сангвинарина из клеточных культур мака путем элиситации алкалоида гомогенатом из мицелия гриба питиум (Pythium aphanidermatum(Edson.) Fitzp.).
Кроме элиситоров грибного и микробного происхождения биосинтез вторичных продуктов могут стимулировать химические вещества. Так, сульфат ванадия способствовал увеличению почти в 2 раза содержания индольных алкалоидов аймалицина и катарантина в культивируемых клетках барвинка розового (Vinca roseaL.). В настоящее время не известен ни механизм воздействия VaSO4 на синтез вторичных продуктов, ни место его аккумуляции. Тем не менее обработка клеток ванадием способствовала сокращению периода роста, увеличению выхода индольных алкалоидов и, в отличие от грибных элиситоров, не требовала выращивания грибов и получения из них элиситора.
Способы выращивания. В большинстве микробных систем продукты, синтезируемые микроорганизмами, продуцируются из клетки непосредственно в среду, в связи с чем эффект обратного ингибирования продуктом реакции очень мал. В культуре ткани, как правило, при увеличении концентрации продукта в клетке выше пороговой, срабатывает эффект обратного ингибирования, вследствие чего дальнейшее накопление вещества прекращается. Данное явление часто обусловливает низкое содержание искомых веществ в клетках культуры. Чтобы избежать этого эффекта, исследователи неоднократно предпринимали попытки удалить продукт реакции из клетки по мере его синтеза. Так, для увеличения проницаемости клеток хинного дерева (Cinchona ledgerianaMoens ex Trimen) в культуре и ускорения выхода внутриклеточных алкалоидов применяли диметилсульфоксид. Однако даже при высокой концентрации сурфактанта высвобождение алкалоидов протекало медленно, и большинство обработанных мембран не восстанавливалось.
Для некоторых суспензионных культур весьма экономичным оказался способ выращивания в виде двухфазной системы: водной фазы (питательная среда с растущими клетками) и нетоксичной липофильной фазы (триглицериды или парафины). В результате липофильные вещества, синтезируемые клетками в процессе их роста, переходят в липофильную фазу. Например, суспензионная культура ромашки росла в двухфазной системе, состоящей из водного раствора питательной среды и нетоксичной липофильной фазы (триглицерид миглиол) в течение 22 дней. В результате жирорастворимые продукты, синтезируемые культурой ткани ромашки, накапливались в фазе миглиола в 60 раз большей концентрации, чем в однофазной системе.
Клеточная дифференциация in vitro. В настоящее время большое число экспериментальных данных свидетельствует о том, что образование и накопление вторичных продуктов в растениях — сложный, пространственно организованный процесс, который часто в той или иной форме включает транспортировку этих соединений на клеточном и субклеточном уровнях. В ряде случаев показано резкое разграничение мест первичного синтеза и накопления алкалоидов. Установлено, что эти процессы могут быть локализованы в пределах одного и того же органа или даже одной и той же ткани, но в различных клетках, что говорит об эпигенетическом контроле процесса пространственного разобщения синтеза и накопления конечного продукта. В частности, люпиновые алкалоиды растений семейства Бобовые синтезируются в зеленых частях растений, а накапливаются в корнях. Тропановые алкалоиды синтезируются в корнях, но транспортируются в стебли.
Читайте также: Как мыть рулонные шторы из ткани день ночь
В культурах тканей растений так же, как и в растениях, накопление вторичных метаболитов часто тесно связано со степенью тканевой дифференциации. В культурах ткани обычно формируются секреторные канальцы, млечники, слизевые клетки, железки или специализированные клетки, где накапливаются конечные продукты, т. е. наблюдается процесс разобщения синтеза и накопления вторичных веществ. Установлено, например, что практически все клетки в каллусе маклейи обладают способностью синтезировать изохинолиновые алкалоиды, однако их накопление осуществлялось лишь в специализированных, так называемых «алкалоидных» клетках. В культуре ткани женьшеня обнаружены секреторные каналы, типичные для растений семейства Аралиевые.
Исследователи, работающие с культурами тканей, отмечают, что с образованием в каллусе морфологических структур (побегов, корней, эмбриоидов) содержание искомых продуктов в культуре увеличивается. Например, культура ткани красавки (Atroра belladonna L.) при недифференцированном росте не продуцирует гиосциамин, а при образовании в каллусе корней начинает синтезировать его. Кардиогликозиды в культуре ткани наперстянки шерстистой синтезировались только с образованием эмбриоидов.
Важным диагностическим признаком культуры ткани раувольфии змеиной являются выделительные образования с различной степенью морфологической специализации: клетки-идиобласты и удлиненные секреторные структуры типа млечников. Исследования показали, что основная масса алкалоидов находится именно в них, и максимальное содержание алкалоидов наблюдается в период увеличения количества млечников. Индукция морфогенеза в культуре ткани мака привела к появлению алкалоидов тебаина и морфина, тогда как недифференцированные каллусные ткани накапливали сангвинарин и допамин. Содержание гликоалкалоидов в культуре ткани паслена дольчатого при переходе к эмбриогенезу увеличивалось более чем в 10 раз и достигало 1 % сухой массы клеток, что близко к содержанию гликоалкалоидов во взрослом растении.
Обнаружена корреляция между ультраструктурной организацией пластид (этиопластов и хлоропластов) и биосинтетической способностью каллусных культур чайного растения, что подчеркивает важную роль хлоропластов в образовании фенольных соединений. Ультраструктура клеточных линий коптиса японского, образующих алкалоиды типа протоберберина, существенно отличалась от клеточных линий, не способных к синтезу алкалоидов: в продуцирующих алкалоиды клетках перед началом синтеза алкалоидов обнаружен больший объем вакуолей, чем в непродуцирующих клетках. Кроме того, в продуцирующих клетках можно было наблюдать многочисленные везикулы, содержащие в цитоплазме электронно-плотный осадок.
Селекция—основа биотехнологического использования культур
Промышленное применение культур тканей ЛР в качестве ЛРС предполагает использование высокопродуктивных и стабильных клонов. Известно, что культивирование клеток in vitro может сопровождаться значительным генетическим разнообразием. Речь идет о так называемой сомаклональной изменчивости, которая возникает при длительном культивировании каллуса. Сомаклональные варианты, сохраняя основные свойства прототипа, часто значительно отличаются от него устойчивостью к вирусам, болезням, экологическим стрессам, а иногда несколько измененной биосинтетической способностью и более высокой продуктивностью, т. е. могут затрагивать хозяйственно ценные признаки. На изменчивости клеток в культуреin vitro основана селекция штаммов, обеспечивающая большой выход ценных продуктов метаболизма.
Для увеличения спектра изменчивости используют обработку мутагенами, а также селективные условия культивирования клеток. Спонтанно возникшие или индуцированные мутанты в популяции отбираются на устойчивость к созданным жестким условиям: высоким концентрациям солей, экстремальным температурам, гербицидам, токсинам и др. В результате многих экспериментов удается отобрать устойчивые линии и получить растения-регенераты из стабильной клеточной линии. Часто успех опыта зависит от методов оценки селекционного материала. Так, для количественной оценки содержания аймалина в культуре ткани раувольфии змеиной разработан простой и быстрый метод, основанный на сравнении интенсивности окраски пятен сока каллусных культур, нанесенных на фильтровальную бумагу и обработанных цветным реактивом, со шкалой стандартов. Метод позволяет исключить неперспективные варианты и сократить число культур, подлежащих окончательной проверке.
Успешным оказался способ селекции, основанный на отборе окрашенных участков культуры ткани. Известно, что окраска тканей растений является наследственным признаком, связанным с их химическим составом. Связь между окраской тканей и их химическим составом была использована в селекции культур тканей, высокопродуктивных по содержанию антоцианидинов. После 12 пересевов содержание антоцианидинов увеличилось в 3 раза. Селекция промышленных штаммов воробейника краснокорневого была осуществлена путем отбора и пересадок красных участков каллуса. Высокоалкалоидные культуры ткани барбариса — продуцента берберина — также были получены в результате клонирования и отбора желтых участков каллуса.
Высокопродуктивные клоны культуры ткани табака были селектированы благодаря полуколичественному методу определения алкалоидов с реактивом Драгендорфа. Ярко-голубую флуоресценцию серпентина в УФ-свете (max= 365 нм) использовали в селекции высокоалкалоидных штаммов суспензионной культуры катарантуса розового. Селекция культур тканей с высоким содержанием изохинолиновых, акридоновых и индольных алкалоидов основана на измерении их флуоресценции.
Важным условием биотехнологического использования культур тканей является их стабильность, гарантирующая стандартность ЛРС. Такие культуры суперпродуценты, как амми зубная (Ammi visnaga[L.] Lam.), раувольфия змеиная (Rauwolfia serpentinaBenth.), воробейник красный (Lithospermum erythrorhizonSieb. et Zucc.), на протяжении многих лет зарекомендовали себя как устойчивые. Но для многих тканевых культур, например барвинка розового (Catharanthus roseus [L.] G. Donf.), единственным путем сохранения высокой продуктивности является регулярный поддерживающий отбор, их нестабильность — серьезное препятствие к промышленному использованию.
Клеточная селекция — одна из наиболее перспективных клеточных технологий для создания сортов не только важнейших сельскохозяйственных, но и ЛР. В настоящее время проводится большое количество исследовательских работ по созданию высокопродуктивных штаммов и растений-регенератов методами гибридизации соматических(неполовых) клеток путем слияния протопластов и генной инженерии. Хотя методы соматической гибридизации и генной инженерии (с введением в ядерный геном генов синтеза фармакологически ценных веществ) еще не получили промышленного развития, однако ученые считают, что за ними будущее и генная инженерия станет естественным приемом в создании новых необходимых человеку ЛР — продуцентов ЛС.
Трансгенными называются растения тех видов, в которых успешно функционируют гены (или ген), пересаженные из растений или животных других видов. Делается это для того, чтобы растение-реципиент получило новые удобные для человека свойства: повышенную устойчивость к вирусам, гербицидам, вредителям и болезням растений. Было обнаружено, что опухолеобразующие Ti-плазмиды агробактерий (Ti — tumor inducing), представляющие миникольцевые ДНК, являются великолепной природной векторной системой, которую в настоящее время используют для переноса генов в растения. Плазмидная тДНК (transferred DNA) обладает двумя свойствами, делающими ее по существу идеальным вектором для введения чужеродных генов в клетки растений. Во-первых, круг хозяев агробактерий очень широк: они трансформируют клетки практически всех двудольных растений (иногда даже однодольных, в том числе злаков). Во-вторых, интегрированная в состав генома растения тДНК наследуется как простой доминантный признак в соответствии с законами Менделя, а ее гены имеют собственные промоторы (регуляторная область гена, определяющая время и место его экспрессии), под контролем которых могут экспрессироваться вставленные в тДНК чужеродные гены. С помощью Ti-плазмид агробактерий уже получены следующие продукты: морозоустойчивая свекла, светящаяся в сумерках газонная трава, устойчивый к колорадскому жуку картофель и даже банан, съедая который, человек получает «прививку от тропических болезней».
Во всем мире активно ведутся работы по созданию на основе трансгенных растений так называемых«съедобных вакцин», которые в дальнейшем можно будет использовать для предупреждения наиболее опасных болезней. Например, ученые Сибирского отделения РАН разрабатывают противотуберкулезную вакцину, для чего используют гены человека, кодирующие синтез специфических антител, и вводят их в геном клеток растений, а их коллеги из Института биофизики и клеточной инженерии НАН Беларуси намерены получить картофель, содержащий иммуноглобулины. В зависимости от условий клетки в культуре in vitro могут делиться анархически, образуя неорганизованную каллусную массу, либо менять программу своего поведения и делиться организованно с образованием зачатков корней, стеблей, зародышей. Из этих зачатков растений в культур еin vitro затем можно регенерировать растения.
Легче всего вызвать морфогенез (образование органов и тканей) и регенерацию растения, используя зародыши и почки, а также стеблевые меристемы, но даже из зародышей, изолированных на ранних стадиях развития, или апикальных меристем стебля очень маленьких размеров для получения растений нужны дополнительные условия, например очень богатые питательные среды. Обычно в каждом случае разрабатывают условия культивирования и соответствующие питательные среды. Стеблевая верхушечная меристема, как правило, свободна от вирусной инфекции, микоплазм и возбудителей других инфекций, поэтому культивирование апикальных меристем, а затем быстрое клональное размножение здоровых растений — основа получения безвирусного посадочного материала растений.
Велико значение культуры тканей для быстрого клонального микроразмножения растений, используя клоны генетически идентичных клеток. Этот процесс, в сравнении с традиционной техникой вегетативного размножения черенками, отводками, усами, прививками, представляется ювелирным. Он идет быстро и с высоким выходом посадочного материала: от материнской клетки за год можно получить 10 5 —10 6 растений. Растения-регенераты затем адаптируются к условиям почвы и переводятся в обычную агрокультуру. Сейчас это основное направление в биотехнологии сельскохозяйственных и ЛР. Технологии клонального микроразмножения — важное дополнение к традиционной селекции растений. Становится возможным быстро размножить уникальный генотип или новый сорт, что сильно ускоряет его практическое использование. В настоящее время найдены условия размножения более 500 экономически важных или исчезающих видов дикорастущих растений и тем самым сохранения природного биоразнообразия, представляющего собой одно из богатств государства и важнейшее условие нормального существования и жизнедеятельности человека. Многие из таких видов размножаются уже в производственных условиях.
Технологии микроклонального размножения ЛР разработаны в отделе биологии клетки и биотехнологии Института физиологии растений АН России (Москва) для мандрагоры туркменской, аристолохии маньчжурской, женьшеня; в Химико-фармацевтической академии (Санкт-Петербург) — для ряда видов раувольфии; в ВИЛАРе — для стефании гладкой. Аналогичные работы проводятся в отделе биохимии и биотехнологии растений ЦБС НАН Беларуси.
Таким образом, научной основой биотехнологического использования культур тканей растений в фармакогнозии является способность клеток in vitroсинтезировать самые разнообразные группы веществ: гликозиды, фенольные соединения, кардиостероиды, сапонины, лигнаны, флавоноиды, терпеноиды, алкалоиды и другие природные соединения. Все они — звенья сложныхцепей метаболизма у растений, где выполняют определенные, часто весьма важные, функции.
В заключение приведем в качестве примеров используемые в клинической практике ЛС, полученные на основе каллусныхи суспензионныхкультур клеток ЛР: шиконин (для лечения кожныхзаболеваний), дигоксин (для лечения сердечно-сосудистыхзаболеваний), берберин (для лечения кишечныхрастройств — в качестве бактерицидного средства), диосгенин (как противозачаточное средство), панаксозиды (используются в качестве адаптогенов, укрепляющихиммунитет). При производстве настоек женьшеня выращивание этой культуры на плантацияхпо выходу панаксозидов имеет преимущество перед каллусным сырьем, однако препараты, получаемые из каллусного сырья, менее токсичны.
В последние годы число информационныхсообщений, касающихся синтеза вторичныхметаболитов в культурахтканей растений и имеющихзначимость для фармацевтической промышленности, возросло. По-видимому, переход от научныхразработок к промышленному производству ЛС с использованием клеточных культур ЛР только начинается.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом