
В сложных механизмах транспорта газов кровью и газообмена в тканях важная роль отводится эритроцитам, ответственным за доставку О2 к различным органам и удаление образующегося в процессе метаболизма СО2.
Эритроцит – безъядерная клетка, лишенная митохондрий, основным источником энергии для эритроцита служит глюкоза, метаболизируемая в гексозомонофосфатном шунте или цикле Эмбдена-Мейергофа. Транспорт О2 обеспечивается в значительной мере гемоглобином, состоящим из белка глобина и гема. Последний представляет собой комплексное соединение железа и порфирина. Глобин представляет собой тетрамер полипептидной цепи. Hb A (HbA) – основной гемоглобин взрослых содержит 2 – альфа и 2 – бета – цепи, Hb A2 – содержит две альфа и две дельта цепи.
Гем состоит из иона железа, встроенного в порфириновое кольцо. Ион железа гема обратимо связывает одну молекулу О2. С одной молекулой Hb максимально связываются 4 молекулы О2 с образованием оксигемоглобина.
Гем может подвергаться не только оксигенации, но и истинному окислению, когда железо становится из двухвалентного трехвалентным. Окисленный гем носит название гематина, а молекула гемоглобина становится метгемоглобином. В крови человека метгемоглобин находится в незначительных количествах, его уровень резко возрастает при отравлениях. Метгемоглобин не способен отдавать кислород тканям.
В норме метгемоглобин составляет менее 3% общего Hb крови. Основная форма транспорта О2 – в виде оксигемоглобина. Кислород транспортируется артериальной кровью не только в связи с гемоглобином, но и в растворенном виде. Принимая во внимание тот факт, что 1 г Hb может связать 1,34 мл О2, кислородная емкость крови в среднем у взрослого человека составляет около 200 мл/л крови. Одним из показателей кислородного транспорта является насыщение артериальной крови О2(Sa O2), равного отношению О2, связанного с Hb, к кислородной емкости крови:
SaO2=O2, связанного с Hb/O2 емкость крови* 100%.
В соответствии с кривой диссоциации оксигемоглобина насыщение артериальной крови кислородом в среднем составляет 97%, в венозной крови – 75%.
PaO2 в артериальной крови около 100 мм. рт. ст., а в венозной – около 40 мм. рт. ст.
Количество растворенного кислорода в крови пропорционально парциальному давлению О2 и коэффициэнту его растворимости.
Последний для О2 составляет 0,0031/100 мл крови/ 1 мм. рт. ст.. Таким образом, 100 мл крови при PaO2, равном 100 мм. рт. ст., содержит менее 0,31 мл O2.
Диссоциация оксигемоглобина в тканях обусловлена главным образом химическими свойствами гемоглобина, а также рядом других факторов – температурой тела, рН среды, р СО2.
При понижении температуры тела наклон кривой диссоциации оксигемоглобина возрастает, а при ее повышении – снижается, и соответственно снижается сродство Hb к О2.
При снижении рН, т.е. при закислении среды, сродство гемоглобина к О2 уменьшается. Увеличение напряжения в крови СО2 также сопровождается снижением сродства Hb к О2 и уплощением кривой диссоциации оксигемоглобина.
Известно, что степень диссоциации оксигемоглобина определяется содержанием в эритроцитах некоторых фосфорорганических соединений, главным из которых является 2,3 – ДФГ (2,3 дифосфоглицерат), а также содержанием в эритроцитах катионов. В случаях развития алкалозов, поглощение О2 в легких увеличивается, но в то же время затрудняется отдача кислорода тканями. При ацидозах наблюдается обратная картина.
4.2.Утилизация кислорода тканями
Тканевое или клеточное дыхание включает три стадии. На первой стадии пируват, аминокислоты и жирные кислоты окисляются до двухуглеродных фрагментов ацетильных групп, входящих в состав ацетилкофермента А. Последние на втором этапе окисления включаются в цикл лимонной кислоты, где происходит образование высокоэнергетических атомов водорода и высвобождение СО2 – конечного продукта окисления органических субстратов. На третьей стадии клеточного дыхания атомы водорода делятся на протоны (Н+) и «высокоэнергетические» электроны, передающиеся по дыхательной цепи на молекулярный О2 и восстанавливающие его до НО2. Перенос электронов сопряжен с запасом энергии в форме АТФ, т.е. с окислительным фосфорилированием (рис.6).
Касаясь патогенеза метаболических сдвигов, свойственных гипоксическим состояниям, следует отметить, что в организме человека более 90% всего потребляемого кислорода восстанавливается с участием цитохромоксидазы митохондрий, и лишь около 10% кислорода метаболизируется в тканях с участием оксигеназ: диоксигеназы и монооксигеназы.
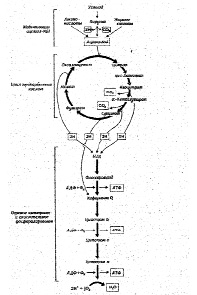
Рис.6. Схема тканевого дыхания. Конечные продукты каждой стадии даны в рамке (Ленинджер А., 1999)
Наиболее многочисленны и сложны монооксигеназные реакции, протекающие в эндоплазматическом ретикулуме клеток при участии цитохрома Р-450 и обеспечивающие гидроксилирование субстрата (стероидных гормонов, лекарственных препаратов и различных др. соединений) и, как правило, его инактивацию.
Диоксигеназы катализируют реакции, в которых в молекулу органического субстрата включаются оба атома молекулы кислорода (например, реакция окисления катехола молекулярным кислородом с раскрытием кольца).
В реакциях, связанных с переносом электронов, т.е. в реакциях окисления-восстановления, где, как указывалось выше, используется более 90% потребляемого кислорода, атомы водорода, отщепленные дегидрогеназами от субстратов в цикле лимонной кислоты, передают свои электроны в цепь переноса электронов и превращаются также в Н +. Как известно, помимо 4 пар атомов водорода, поставляемых каждым оборотом цикла лимонной кислоты, образуются и другие атомы водорода, отщепленные дегидрогеназами от пирувата, жирных кислот и аминокислот в процессе их расщепления до Ацетил-СоА и других продуктов.
Таким образом, все атомы водорода, отщепляемые дегидрогеназами от субстратов, передают свои электроны в дыхательную цепь к конечному акцептору электронов – кислороду.
Касаясь последовательности транспорта электронов в окислительно-восстановительных реакциях, протекающих на внутренней мембране митохондрий, прежде всего, следует отметить, что от всех НАД – зависимых реакций дегидрирования восстановленные эквиваленты переходят к митохондриальной НАДН – дегидрогеназе, затем через ряд железосерных ферментов передаются на убихинон М цитохрому b. Далее электроны переходят последовательно на цитохромы С1 и С, затем на цитохромы аа 3 (цитохромоксидазу – медьсодержащий фермент). В свою очередь цитохромоксидаза передает электроны на кислород. Для того, чтобы полностью восстановить кислород с образованием 2-х молекул воды требуется 4 электрона и четыре Н+ .
Скорость утилизации О2 в различных тканях различна. В среднем взрослый человек потребляет 250 мл О2 в 1 мин. Максимальное извлечение О2 из притекающей артериальной крови свойственно миокарду.
Кислород используется в клетках, в основном в метаболизме белков, жиров, углеводов, ксенобиотиков, в окислительно-восстановительных реакциях в различных субклеточных фракциях: в митохондриях, в эндоплазматическом ретикулуме, в реакциях липопероксидации, а также в межклеточном матриксе и в биологических жидкостях.
Читайте также: За 38 м ткани двух сортов уплатили 104 р
Коэффициент утилизации О2 в тканях равен отношению потребления О2 к интенсивности его доставки, широко варьирует в различных органах и тканях.
В условиях нормы минимальную потребность в О2 проявляют почки и селезенка, а максимальную потребность – кора головного мозга, миокард и скелетные мышцы, где коэффициент утилизации О2 колеблется от 0,4 до 0,6, а в миокарде до 0,7. При крайне интенсивной физической работе коэффициент утилизации О2 мышцами и миокардом может возрастать до 0,9.
Обмен дыхательных газов в тканях происходит в процессе свободной и облегченной диффузии. При этом О2 переносится по градиенту напряжения газа из эритроцитов и плазмы крови в окружающие ткани.
Одновременно происходит диффузия СО2 из тканей в кровь. На выход О2 из крови в ткани влияет диссоциация оксигемоглобина в эритроцитах, что обеспечивает так называемую облегченную диффузию О2. Интенсивность диффузионного потока О2 и СО2 определяется градиентом их напряжения между кровью и тканями, а также площадью газообмена, плотностью капилляров, распределением кровотока в микроциркуляторном русле. Интенсивность окислительных процессов в тканях определяется величиной критического напряжения О2 в митохондриях, которое в условиях нормы должно превосходить 0,1-1 мм рт. ст.
Соответствие доставки О2 к органам и тканям, возросшим потребностям в оксигенации обеспечивается на клеточном, органном уровнях за счет образования метаболитов изнашивания, а также при участии нервных, гормональных и гуморальных влияний.
Основная масса углекислого газа (СО2) образуется в организме как конечный продукт различных метаболических реакций и транспортируется к легким с кровью. Вдыхаемый воздух содержит лишь незначительное количество СО2.
Транспорт СО2 кровью осуществляется в 3-х состояниях: в виде аниона бикарбоната, в растворенной форме и в виде карбаминовых соединений.
СО2 хорошо растворяется в плазме крови и в артериальной крови, около 5% от общей двуокиси углерода содержится в крови в растворенной форме.
Анион бикарбоната составляет около 90% от общего содержания СО2 в артериальной крови: СО2+Н2О — Н++НСО-3.
Эта реакция медленно протекает в плазме крови, но чрезвычайно интенсивно происходит в эритроцитах при участии фермента карбоангидразы. Мембрана эритроцита относительно непроницаема для Н+, как и вообще для катионов, но в тоже время, проницаема для ионов НСО-3, выход которых из эритроцитов в плазму обеспечивается притоком Cl- из плазмы в эритроциты. Часть Н+ забуферивается гемоглобином с образованием восстановленного гемоглобина.
Третьей формой транспорта СО2 кровью являются карбаминовые соединения, образованные взаимодействием СО2 с концевыми группами белков крови преимущественно с гемоглобином:
Hb NH2 + CO2 — Hb NH COOH > Hb NH COO + Н+
Карбаминовые соединения составляют около 5% от общего количества СО2, транспортируемого кровью.
В оксигенированной артериальной крови напряжение СО2 составляет 40 мм. рт. ст., а в венозной крови Рv СО2 равно 46 мм. рт. ст.
4.4.Связывание гемоглобина с окисью углерода
Угарный газ (СО) – окись углерода обладает значительно большим сродством к гемоглобину, чем О2, с последующим образованием карбоксигемоглобина. СО входит в состав бытового газа, а также выделяется при работе двигателей внутреннего сгорания. При концентрации СО во вдыхаемом воздухе всего в количестве 7*10- 4 около 50% гемоглобина превращается в карбоксигемоглобин. В норме в крови содержится около 1% HbCO, у курильщиков – 3% . В крови водителей такси концентрация карбоксигемоглобина достигает 20%. Карбоксигемоглобин диссоциирует с отдачей О2 в 200 раз медленней оксигемоглобина и в то же время препятствует его диссоциации в тканях.
Газообмен в тканях
В тканях кровь отдает кислород и поглощает углекислоту. Газообмен в капиллярах тканей большого круга, так же как и в легочных капиллярах, обусловлен диффузией вследствие разности парциальных напряжений газов в крови и в тканях.
Напряжение углекислого газа в клетках может достигать 60 мм, в тканевой жидкости оно весьма изменчиво и в среднем составляет 46 мм, а в притекающей к тканям артериальной крови — 40 мм рт. ст. Диффунтируя по направлению более низкого напряжения, углекислый газ переходит из клеток в тканевую жидкость и далее в кровь, делая ее венозной. Напряжение углекислого газа в крови при прохождении ее по капиллярам становится равным напряжению углекислого газа в тканевой жидкости.
Клетки весьма энергично потребляют кислород, поэтому его парциальное напряжение в протоплазме клеток очень низко, а при усилении их активности может быть равно нулю. В тканевой жидкости напряжение кислорода колеблется между 20 и 40 мм. Вследствие этого кислород непрерывно поступает из артериальной крови, приносимой к капиллярам большого круга кровообращения (здесь напряжение кислорода равно 100 мм рт. ст.), в тканевую жидкость. В результате в оттекающей от тканей венозной крови напряжение кислорода значительно ниже, чем в артериальной, составляя 40 мм.
Кровь, проходя по капиллярам большого круга, отдает не весь свой кислород. Артериальная кровь содержит около 20 об.% кислорода, венозная же кровь — примерно 12 об. % кислорода. Таким образом, из 20 об. % кислорода ткани получают 8 об. %, т. е. 40% всего кислорода, содержащегося в крови.
То количество кислорода в процентах от общего содержания его в артериальной крови, которое получают ткани, носит название коэффициента утилизации кислорода. Его вычисляют путем определения разности содержания кислорода в артериальной и венозной крови. Эту разность делят на содержание кислорода в артериальной крови и умножают на 100.
Коэффициент утилизации кислорода меняется в зависимости от ряда физиологических условий. В покое организма он равен 30—40%. При тяжёлой же мышечной работе содержание кислорода в оттекающей от мышц венозной крови уменьшается до 8—10 об. % и, следовательно, утилизация кислорода повышается до 50—60%.
Более быстрый переход кислорода в ткани обеспечивается раскрытием нефункционировавших капилляров в работающей ткани. Повышению коэффициента утилизации способствует также усиленное образование кислот — молочной и угольной, что понижает сродство гемоглобина к кислороду и обеспечивает более быструю диффузию кислорода из крови. Наконец, увеличению утилизации кислорода содействуют повышение температуры работающих мышц и усиление ферментативных и энергетических процессов, протекающих в клетках. Таким образом, доставка кислорода к тканям регулируется в соответствии с интенсивностью окислительных процессов.
Читайте также: Код тн вэд кремнеземная ткань
Биохимические основы работы дыхательной системы

Биохимические основы работы дыхательной системы.
(Глава из учебного пособия «Дыхательная система», Благовещенск, 2011г.)
Роль кислорода у живых организмов.
Любой современный человек, даже весьма далекий от биологии и медицины, без колебания скажет, что роль легких в организме человека и животных связана с поглощением кислорода воздуха и выведением из организма углекислоты. Значительно меньше людей знают что-либо о процессах использования кислорода тканями, несмотря на то, что именно эти процессы раскрывают роль кислорода, как окислителя, выбранного живой природой для извлечения энергии из молекул органических веществ.
Единственным источником энергии для организмов аэробов является окисление поступающих с пищей углеводов, жиров и белков. Этот процесс может протекать и в отсутствие кислорода, т. е. в анаэробных условиях. Однако, анаэробный распад энергетически малоэффективен, поскольку его продукты еще не полностью окислены и в их молекулах сохраняется большое количество энергии. В частности, при анаэробном распаде глюкозы, завершающемся образованием молочной кислоты, количество выделяющейся энергии позволяет запасти 2 моля АТФ, а аэробный распад глюкозы до полностью окисленных продуктов СО2 и Н2О позволяет синтезировать 36 молей АТФ на моль глюкозы.
В отличие от реакции горения в неживой природе, например сгорания входящих в состав бензина углеводородов в двигателе автомобиля, при аэробном распаде веществ у живых организмов кислород не включается в состав окисляемых органических молекул. Ни одна из реакций распада ацетил КоА в цикле Кребса не сопряжена с поглощением кислорода, несмотря на то, что в условиях дефицита кислорода работа цикла замедляется. Для чего же нужен кислорода при аэробном распаде веществ, если он не взаимодействует непосредственно с окисляемыми молекулами? В живой клетке кислород используется для окисления атомов водорода, подхваченных коферментами дегидрогеназ (НАД+, ФАД) от метаболитов сахаров, жирных кислот и аминокислот. Окисление водорода молекулярным кислородом протекает в дыхательной цепи внутренней мембраны митохондрий и представляет оксидазный путь утилизации кислорода в клетке. Оксидазный путь имеет энергетическое значение поскольку выделяющаяся при окислении водорода в дыхательной цепи энергия запасается в ходе окислительного фосфорилирования в виде богатых энергией фосфатных связей АТФ и используется клеткой для различных проявлений жизнедеятельности. Таким образом, в многостадийном процессе утилизации кислорода живыми организмами легкие участвуют на начальном этапе, поглощая кислород из атмосферного воздуха, и на конечной этапе выводя из организма углекислоту, являющуюся продуктом протекающего в тканях окислительного распада органических молекул.
Дыхательная функция легких, связанная с газообменом между атмосферным воздухом и кровью, без сомнения, важнейшая, но далеко не единственная. Важная функция легких состоит в поддержании постоянство состава внутренней среды организма. В частности, легкие обезвреживают попадающие с вдыхаемым воздухом чужеродные для организма вещества – ксенобиотики, участвуют в реакциях клеточного иммунитета и в поддержании постоянства кислотно-основного состояния. В легких синтезируются вещества, необходимых для функционирования самих легких (компоненты системы сурфактанта) и биологически активные вещества, обладающие регуляторным действием на чувствительные к ним клетки различных органов и тканей. Легкие захватывают из крови и разрушают или модифицируют вазоактивные гормоны. Указанные функции называют нереспираторными функциями легких. Рассмотрению нереспираторных функций и особенностей обмена веществ в легких посвящена настоящая глава. Процессы же транспорта газов в легких подробно изложены в главе «Биофизические основы дыхания».
Особенности обмена веществ в легких.
В любом органе, любой ткани протекают процессы обмена веществ. Основные пути метаболизма к настоящему времени достаточно полно выяснены. В обмене веществ в разных органах и тканях есть как общие, так и специфические черты. Процессы распада веществ (катаболизма) обеспечивают выделение энергии из органических молекул и ее запасание в виде веществ-макроэргов или градиентов ионов на клеточных мембранах. С точки зрения сопутствующих химическим реакциям изменениям энергии процессы катаболизма это экзергонические процессы. В экзергонических процессах энергия выделяется. В ходе процессов синтеза (анаболизма) образуются необходимые клеткам и организму в целом вещества. Синтез сложных веществ из более простых (анаболизм) сопряжен с поглощением энергии. Такие процессы являются эндергоническими. Анаболические реакции обеспечивают клетки веществами, необходимыми для поддержания их структуры и функциональной активности. Некоторые из синтезируемых в клетках веществ секретируются наружу и выполняют свою роль вдалеке от места образования.
Долгое время сведения об особенностях метаболизма в легких были весьма ограничены. Причины отставания в развитии наших представлений о роли легких в метаболизме связаны с особенностями их строения. Легкие занимают большую часть грудной клетки, и в то же время масса легких составляет лишь 1% от массы целого организма. Легкие – единственный орган, через который проходит весь объем циркулирующей крови за один кругооборот. Около 30 % массы легких приходится на содержащуюся в них кровь. Следовательно, масса легких пренебрежительно мала по отношению к объему протекающей через них крови. Поэтому исследование разницы в концентрациях метаболитов в притекающей к легким крови и в крови, оттекающей от легких (артерио-венозная разница) мало что дает для понимания биохимических процессов в этом органе поскольку различия крайне незначительны. Исключением является разница в газовом составе крови, но она характеризует не метаболизм легких, а их роль в газообмене.
Для исследования метаболизма легких потребовалось создание экспериментальных моделей in vitro. Примером такой модели является модель изолированного перфузируемого легкого (рисунок 1).
Рисунок 1. Модель изолированных легких. Одна канюля соединена с легочной артерией, вторая с легочной веной. Перфузируемая жидкость циркулирует через легкие.
Перфузия органа искусственной средой позволяет добиться полной отмывки от крови. Поэтому разница в содержании веществ в притекающей и вытекающей из легких жидкости всецело обусловлена биохимическими превращениями этих веществ в клетках легких. Другими моделями для исследования процессов метаболизма в легочной ткани являются срезы ткани легкого, фракции субклеточных органелл и культуры клеток легких. Использование культуры изолированных альвеолоцитов IIтипа позволила детально охарактеризовать роль этих клеток в продукции сурфактанта.
Потребление кислорода легкими
Зная скорость потребления кислорода тем или иным органом, можно сразу же судить об интенсивности обмена веществ в этом органе, и в целом об интенсивности его функционирования. Метаболически активные органы потребляют значительные количества кислорода, а неактивные органы, напротив потребляют значительно меньше кислорода. Через легкие в кровь поступает большое количество кислорода. Однако, на протяжении долгого времени оставалось неясно – сколько кислорода утилизируется самими легкими? Ответ на этот вопрос дали результаты экспериментов с перфузируемыми срезами ткани легких. Скорость поглощения ими кислорода составляет 30-150 мкл О2 мин-1 г-1 сухого веса ткани, что существенно ниже, чем у метаболически активных органов, таких как сердце, почки, головной мозг, щитовидная железа. В то же время, эти величины выше, чем у скелетных мышц в состоянии покоя и у метаболически менее активных органов. Таким образом, скорость потребления кислорода легкими можно расценивать как среднюю. Это согласуется с тем, что на легкие приходится 1% массы организма и легкие утилизируют 1% поступающего в организм кислорода. Следует иметь в виду, что ткань легких неоднородна и представлена различными типами клеток, которые могут существенно отличаться по потреблению О2. В частности, высокое потребление кислорода характерно для альвеолоцитов II типа.
Читайте также: Мрт мягких тканей ротовой полости
Какие субстраты использую легкие в качестве топлива?
В физиологических условиях главным видом клеточного топлива для большинства органов является глюкоза. Ткани, использующие в качестве топлива только глюкозу, называются облигатными потребителями глюкозы. Примером таких тканей являются нервная ткань, эпителий кишечника, мозговое вещество почек, эритроциты. Большинство тканей может использовать в качестве источников энергии наряду с глюкозой и другие субстраты – жирные кислоты, кетоновые тела, аминокислоты. Использование этих субстратов увеличивается в условиях, когда клеткам не хватаем глюкозы — при голодании, усиленной физической работе, в холодных климатических условиях, при некоторых гормональных расстройствах (сахарный диабет). Такие ткани называют факультативными потребителями глюкозы.
К факультативным потребителям глюкозы относятся и легкие, для которых глюкоза является главным энергетическим субстратом для легких в обычных условиях. В опытах с перфузируемыми легкими скорость утилизации глюкозы составляется 40мкмоль ч-1г-1 сухого веса ткани в условиях аэробиоза. При переходе на неэффективный анаэробный метаболизм поглощение глюкозы легкими возрастает более чем в 2 раза.
Из всех органов организма легкие наиболее интенсивно снабжаемый кислородом орган. В этой связи несколько неожиданной представляется высокая интенсивность анаэробного гликолиза в ткани легких. На рисунке 2 представлена «судьба» атомов
углерода глюкозы, метаболизируемой перфузируемыми легкими.
Рисунок 2. Включение атомов углерода глюкозы в изолированном легком
Около половины метаболизируемой глюкозы превращается в лактат, свидетельствуя о важной роли гликолиза и ограниченной роли цикла Кребса.. Лишь 22% глюкозы подвергается в легких аэробному распаду до СО2 и H2O. Их этого количества 1/4 часть углекислоты образуется в цитоплазме в реакциях пентозного цикла и 3/4 в митохондриях в реакциях окислительного декарбоксилирования изоцитрата и альфа-кетоглутарата в цикле трикарбоновых кислот. Возможное объяснение высокой интенсивности гликолиза в легких состоит в том, что ткань легких сильно гетерогенна по клеточному составу и во многих типах клеток количество митохондрий, в которых протекают реакции цикла Кребса, невелико.
Довольно значительная часть атомов углерода глюкозы включаются в состав белков и нуклеиновых кислот (18%).Небольшая часть глюкозы превращается в жирные кислоты (4%), другие липиды (3%) и полисахариды (гликоген) (5%).
Наряду с глюкозой легкие способны окислять жирные кислоты.
Вклад аэробного метаболизма в энергообеспечение легких
Описанные выше результаты свидетельствуют, что большая часть глюкозы в легких подвергается анаэробному распаду с образованием лактата. Из этого можно было бы предполагать, что вклад аэробного метаболизма в энергообеспечение легких должен быть не очень существенным. Однако, результаты экспериментов с влиянием игибиторов аэробного метаболизма и разобщителей дыхания и фосфорилирования в митохондриях на содержание АТФ и отношение АТФ/АДФ в перфузируемых легких не подтвержают подобное заключение и, напротив, однозначно свидетельствуют о важной роли аэробного метаболизма в энергообеспечении жизнедеятельности легких (рисунок 3).
В перфузируемых изолированных легких содержание АТФ (
10 мкмоль/г сухого веса ткани) аналогично таковому во многих тканях, функционирующих за счет аэробного метаболизма, а отношение АТФ/АДФ составляет 8,5. В присутствии ингибитора дыхания СО содержание АТФ и отношение АТФ/АДФ в легких уменьшаются в 2-3 раза. Воздействие разобщителем дыхания и фосфорилирования динитрофенолом также приводит к уменьшению содержания АТФ и индекса АТФ/АДФ в ткани легкого, хотя и в меньшей степени, чем при действии СО.
Рисунок 3. Влияние СО и динитрофенола на содержание АТФ (слева)
и отношение АТФ/АДФ (справа) в изолированном легком.
Энергозависимые процессы в легких
В легких не протекают энергозависимые процессы, подобные мышечному сокращению, всасыванию веществ в эпителии кишечника или почечных канальцев, проведению нервного импульса. Главная функция легких – газотранспортная не требует энергообеспечения. В главе «Биофизические основы дыхания» говорилось, что перенос газов между альвеолярным воздухом и кровью представляет из себя простой диффузионно-контролируемый процесс. Тем не менее, ряд протекающих в легких процессов, такие как цилиарный транспорт в бронхах, фагоцитарная активность альвеолярных макрофагов, сокращение гладкомышечных элементов бронхов, синтез главного компонента сурфактанта — дипальмитоилфосфатидилхолина и его секреция в просвет альвеол требуют интенсивного энергообеспечения и замедляются в условиях гипоксии.
Сурфактант — поверхностно-активное вещество липопротеидной природы, находящееся на границе вода-воздух, выстилающее изнутри альвеолы и препятствующее спадению альвеол при выдохе за счет снижения поверхностного натяжения. Сурфактант секретируется специальной разновидностью клеток легких — альвеолоцитами II типа. Врожденный дефицит сурфактанта проявляется нераскрытием легкого у новорожденных — респираторным дистресс-синдром (РДС).
Главными компонентами сурфактанта являются фосфолипиды, белки и отчасти полисахариды. В таблице 1 приведены сведения о химическом составе сурфактанта.
Липиды
Липиды составляют более 90% массы сурфактанта и половина этого количества приходится на насыщенный глицерофосфолипид – дипальмитоилфосфатидилхолин. Вторым компонентом сурфактанта является другой глицерофосфолипид – фосфатидилглицерин, в состав которого входят остатки преимущественно ненасыщенных жирных кислот. Фосфолипиды сурфактанта образуют поверхностный монослой на внутренней стенке альвеол (рисунок 4). Благодаря ориентации полярных головок фосфолипидов в водную фазу, а гидрофобных жирнокислотные хвостов в воздух, происходит уменьшение поверхностного натяжения на границе раздела вода-воздух. В
Таблица 1. Основые компоненты сурфактанта
Содержание индивидуальных фосфолипидов (%) и индивидуальных белков в условных единицах
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом