
Процесс возбуждения посредством нервных импульсов органов тканей нервных центров
Неврология. Общие данные. Нейрон. Нейроцит. Синапс.
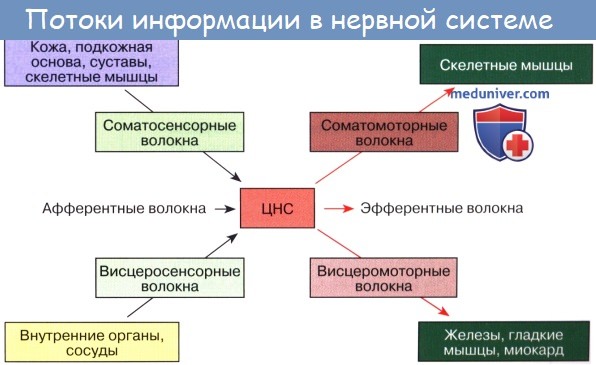
Одним из основных свойств живого вещества является раздражимость. Каждый живой организм получает раздражения из окружающего его мира и отвечает на них соответствующими реакциями, которые связывают организм с внешней средой. Протекающий в самом организме обмен веществ в свою очередь обусловливает ряд раздражений, на которые организм также реагирует. Связь между участком, на который падает раздражение, и реагирующим органом в высшем многоклеточном организме осуществляется нервной системой.
Проникая своими разветвлениями во все органы и ткани, нервная система связывает все части организма в единое целое, осуществляя его объединение, интеграцию.
Следовательно, нервная система есть «невыразимо сложнейший и тончайший инструмент сношений, связи многочисленных частей организма между собой и организма как сложнейшей системы с бесконечным числом внешних влияний» (И. П. Павлов).
В основе деятельности нервной системы лежит рефлекс (И. М. Сеченов). «Это значит, что в тот или иной рецепторный (воспринимающий. — М. П.) нервный прибор ударяет тот или иной агент внешнего или внутреннего мира организма. Этот удар трансформируется в нервный процесс, в явление нервного возбуждения. Возбуждение по нервным волокнам, как по проводам, бежит в центральную нервную систему и оттуда благодаря установленным связям по другим проводам приносится к рабочему органу, трансформируясь, в свою очередь, в специфический процесс клеток этого органа» (И. П. Павлов).
Основным анатомическим элементом нервной системы является нервная клетка, которая вместе со всеми отходящими от нее отростками носит название нейрона, или нейроцита. От тела клетки отходят в одну сторону один длинный (осевоцилиндрический) отросток — аксон, или нейрит, в другую сторону — короткие ветвящиеся отростки — дендриты.
Передача нервного возбуждения внутри нейрона идет в направлении от дендритов к телу клетки от нее к аксону; аксоны проводят возбуждение в направлении от тела клетки. Передача нервного импульса с одного нейрона на другой осуществляется посредством особым образом построенных концевых аппаратов, или синапсов (от греч. synapsis — соединение). Различают аксосоматические связи нейронов, при которых разветвления одного нейрона подходят к телу клетки другого нейрона, и филогенетически более новые аксодендритические связи, когда контакт осуществляется с дендритами нервных клеток.
14. Особенности распространения возбуждения в цнс
Особенности распространения возбуждения в ЦНС в основном определяются свойствами нервных центров:
1. ОДНОСТОРОННЕЕ ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ. В ЦНС возбуждение может распространяться только в одном направлении: от ре-цепторного нейрона через вставочный к эфферентному нейрону, что обусловлено наличием синапсов.
2. БОЛЕЕ МЕДЛЕННОЕ ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ ПО СРАВНЕНИЮ С НЕРВНЫМИ ВОЛОКНАМИ. Промежуток времени от момента нанесения раздражения на рецептор до ответной реакции исполнительного органа называется временем рефлекса. Большая его часть тратится на проведение возбуждения в нервных центрах, где возбуждение проходит через синапсы. На выделение и диффузию медиатора в синапсе требуется промежуток времени в 1,5-2 мс (синоптическая задержка). Чем больше нейронов в рефлекторной дуге, тем продолжительнее время рефлекса.
3. СУММАЦИЯ ВОЗБУЖДЕНИЙ (ИЛИ ТОРМОЖЕНИЯ). Нервные центры могут суммировать афферентные импульсы, что проявляется в усилении рефлекса при увеличении частоты раздражений или числа раздражаемых рецепторов. Различают два вида суммации: временная суммация — если импульсы приходят к нейрону по одному и тому же пути через один синапс с коротким интервалом; пространственная суммация связана с суммированием ВПСП, возникающих одновременно в разных синапсах одного нейрона.
4. КОНВЕРГЕНЦИЯ. В нервном центре несколько клеток могут передавать импульсы к одному нейрону, т. е. возбуждения конвергируют на нем. Конвергенция может быть результатом прихода возбуждающих или тормозных входных сигналов ог различных источников.
5. ДИВЕРГЕНЦИЯ И ИРРАДИАЦИЯ. Возбуждение даже единственного нервного волокна, по которому импульсы поступают в нервный центр, может послужить причиной возбуждения множества выходящих из центра нервных волокон. Морфологическим субстратом широкого распространения импульсов (иррадиации) возбуждения является ветвление аксонов и наличие большого числа вставочных нейронов в пределах центра.
15. Физиология автономной (вегетативной) нервной системы
ВНС (вегетативная нервная система) приспосабливает работу внутренних органов к изменениям окружающей среды. ВНС обеспечивает гомеостаз (постоянство внутренней среды организма). ВНС также участвует во многих поведенческих актах, осуществляемых под управлением головного мозга, влияя не только на физическую, но и на психическую деятельность человека.
ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА — часть нервной системы, обеспечивающая деятельность внутренних органов, регуляцию сосудистого тонуса. Взаимодействуя с соматической нервной системой и эндокринной системой, она обеспечивает поддержание постоянства гомеостаза и адаптацию в меняющихся условиях внешней среды.
Это автономная нервная система, активность которой не контролируется нашим сознанием. Поэтому мы не можем по своему желанию остановить собственное сердце или прекратить процесс переваривания пищи в желудке. Под контролем этой системы находится активность различных желез, сокращение гладких мышц, работа почек, сокращение сердца и многие другие функции. Вегетативная нервная система поддерживает на заданном природой уровне кровяное давление, потоотделение, температуру тела, обменные процессы, деятельность внутренних органов, кровеносных и лимфатических сосудов. Вместе с эндокринной системой она регулирует постоянство состава крови, лимфы, тканевой жидкости (внутренней среды) в организме, управляет обменом веществ и осуществляет взаимодействие отдельных органов в системах органов (дыхания, кровообращения, пищеварения, выделения и размножения).
Вегетативная нервная система состоит из двух отделов: СИМПАТИЧЕСКОГО И ПАРАСИМПАТИЧЕСКОГО, функции которых, как правило, противоположны.
Если нервы симпатического отдела стимулируют какую-то реакцию, то нервы парасимпатического ее подавляют. Эти процессы разнонаправленного воздействия в конечном итоге взаимно уравновешивают друг друга, в результате функция поддерживается на соответствующем уровне. Действие лекарств часто направлено именно на возбуждение или торможение одного из таких противоположных по своей направленности влияний.
Возбуждение симпатических нервов вызывает расширение сосудов головного мозга, кожи, периферических сосудов; расширение зрачка; снижение выделительной функции слюнных желез и усиление — потовых; расширение бронхов; ускорение и усиление сердечных сокращений; сокращение мышц, поднимающих волос; ослабление моторики желудка и кишечника; усиление секреции гормонов надпочечников; расслабление мочевого пузыря и оказывает возбуждающее действие на половые органы, также вызывая сокращение матки.
По парасимпатическим нервным волокнам отдаются «приказы», обратные по своей направленности: например, сосудам и зрачку — сузиться, мускулатуре мочевого пузыря — сократиться и так далее.
Вегетативная нервная система очень чувствительна к эмоциональному воздействию. Печаль, гнев, тревога, страх, апатия, половое возбуждение — эти чувства вызывают изменения функций органов, находящихся под контролем вегетативной нервной системы. Например, внезапный испуг заставляет сильнее биться сердце, дыхание становится более частым и глубоким, в кровь из печени выбрасывается глюкоза, прекращается выделение пищеварительного сока, появляется сухость во рту.
В нашем теле все внутренние ткани и органы, «подчиненные» вегетативной нервной системе, снабжены нервами, которые как датчики собирают информацию о состоянии организма и передают ее в соответствующие центры, а от них доносят до периферии корректирующие воздействия.
Так же как и центральная нервная система, вегетативная система имеет чувствительные (афферентные) окончания (входы), обеспечивающие возникновение ощущений, и исполнительные (двигательные, или эфферентные) окончания, которые передают из центра модифицирующие воздействия к исполнительному органу. Физиологически этот процесс выражается в чередовании процессов возбуждения и торможения, в ходе которых происходит передача нервных импульсов, возникающих в клетках нервной системы (нейронах).
На основании структурно-функциональных свойств автономную нервную систему принято делить на симпатическую, парасимпатическую и метасимпатическую части.
Как протекают процессы возбуждения и торможения в нервной системе?
Деятельность нервной системы основана на процессах возбуждения и торможения, находящихся между собой в постоянно изменяющихся соотношениях. Эти процессы вызываются внешней или внутренней средой, которая воздействуют на нервные клетки-нейроны и вызывает раздражение.
В свою очередь организм животного и человека обладает свойством раздражимости. Раздражимость — это способность внутриклеточных образований, клеток, тканей и органов тела реагировать изменением структур и функций на воздействия факторов внешней и внутренней среды.
Нейрон принимает сигналы от рецепторов и других нейронов, перерабатывает их и в форме нервных импульсов передает к центростремительным нервным окончаниям. Когда этих сигналов нет, он находится в состоянии покоя. В нейроне, находящемся в состоянии покоя, протекают электрохимические процессы, обеспечивающие равновесие между нейроном и внешней средой.
Возбуждение — процесс высвобождения нейроном собственной энергии в ответ на раздражение, ведущий к генерализации потенциалов действия и распространению импульсной активности в нервной системе. В нейроне, находящемся в состоянии возбуждения, нарушается равновесие внутренних электрохимических процессов, что приводит к его активному ответу на воздействия внешней среды. Передача возбуждения от нейрона к нейрону осуществляется с помощью двух механизмов: 1) индукционного, благодаря влиянию электрических полей возбужденных нервных клеток на соседние; 2) путем передачи возбуждения нервных клеток через определенные соединения синапсов. Распространение возбуждения происходит диффузно (во все стороны) или направленно в зависимости от состояния окружающих нейронов.
Торможение — активный процесс, в результате которого возбуждение нейрона прекращается или затрудняется его возникновение. Он проявляется в ослаблении или прекращении деятельности, специфической для данной системы организма. В нейроне, находящемся в состоянии торможения, как правило, начинается восстановление равновесия происходящих в нем электрохимических процессов.
Возбуждение и торможение не остаются в том месте нервной системы, где они возникли, а распространяются на другие участки и отделы, с тем чтобы потом вернуться и сосредоточиться в зоне первоначального возникновения. Кроме того, возникновение одного из процессов вызывает развитие другого. Очаг возбуждения, возникший
в коре головного мозга, воздействуя на соседние участки, вызывает там торможение. Может быть и так: возникшее в одном очаге нервной системы возбуждение переходит затем в торможение, Установлено, что в обычной жизни раздражители возникают не в виде изолированных, единичных явлений, а как комплекс всевозможных раздражителей. Кроме того, организм отвечает не одной изолированной реакцией, а комплексной. Нервной системе, таким образом, присуща системная деятельность.
Проведение возбуждения по нервному волокну
Механизм проведения возбуждения в нервных волокнах объясняется возникновением локальных токов между возбужденными и невозбужденными участками нервного волокна. При возбуждении активируются натриевые каналы, Потенциал действия возникает на аксоном холмике нейрона в результате активации натриевых каналов, происходит перезарядка мембраны нейрона — отрицательный заряд меняется на положительный. Мембрана аксона на всем
протяжении омывается солевым раствором, который хорошо проводит ток. Поскольку рядом с деполяризованным, заряженным положительно участком мембраны расположен поляризованный участок, заряженный отрицательно, возникает разность потенциалов, способная деполяризовать мембрану до КУД, поэтому и на соседнем участке возникает ПД (рис. 16).

Рисунок 16. Проведение возбуждения по нервному волокну
В безмиелиновых волокнах возбуждение распространяется непрерывно, а в миелинизированных — «скачками» между перехватами Ранвье (сальтаторный тип проведения). Дело в том, что миелин оболочки делает ее не проницаемой для электрического тока, поэтому ее сопротивление почти в 1000 раз больше, чем в перехватах. Следовательно, деполяризация до КУД и развитие ПД может возникнуть только в местах перехватов – нервный импульс делает «скачок» сразу на большое расстояние и скорость его распространения существенно увеличивается (рис 17). Такое проведение возбуждения называется сальтаторное.


Сальтаторное проведение импульса по миелинизированному волокну
Закономерности проведения возбуждения по нервному волокну:
1. Возбуждение может распространяться в любом направлении, потому что соседние участки с обеих сторон электроотрицательны по отношению к возбужденному участку.
2. Возбуждение распространяется не затухая, т.к. разность потенциалов деполяризует участок мембраны до КУД, следовательно, возникает ПД стандартной величины.
3. Скорость проведения возбуждения тем больше, чем выше амплитуда потенциала действия, потому что больше возникающая на соседних участках аксона разность потенциалов.
4. Скорость проведения прямо пропорциональна диаметру нервного волокна.
5. Частота импульсов не изменяется.
6. Возбуждение проводится изолированно по каждому нервному волокну.
Проведение возбуждения в синапсах.
Передача нервного импульса с нервного волокна на другую клетку осуществляется посредством СИНАПСОВ. Синапс — специализированная зона контакта между аксоном и другим нейроном, мышечной или секреторной клеткой, обеспечивающая передачу нервного импульса с нервного волокна на эффекторную клетку.

Рисунок 18. Зоны синаптических контактов отмечены стрелками
Существуют синапсы с электрическим и химическим способами передачи. Электрических синапсов в организме человека немного, плотные контакты между клетками обеспечивают такой же вариант передачи импульса, как и в нервных волокнах – с помощью возникающих в месте контакта местных токов. Мы рассмотрим только химические синапсы.
В химических синапсах возбуждение передается с помощью химического вещества — МЕДИАТОРА.
В структуре синапса принято выделять пресинаптическую мембрану, образованную терминалью аксона, синаптическую щель и постсинаптическую мембрану, каждая структура выполняет свою функцию (рис.19).

Рисунок 19. Структура синапса.
Пресинаптическаямембрана терминали аксона представлена, как правило, утолщением, в котором содержится множество гранул диаметром от 20 — 40 до 120 нм. С помощью гистохимических методов было установлено, что в гранулах содержится химические вещества, различные для различных нейронов, эти химические вещества были названымедиаторами– химическими «посредниками» в передаче электрического импульса с пресинаптической к постсинаптической мембране.
— биогенными аминами: норадреналин, дофамин, гистамин, серотонин;
— аминокислотами: глутаминовая кислота, глицин;
— олигопептидами: вещество Р, энкефалины, эндорфины;
— метаболитами: аденозин, АТФ, АДФ;
— широко распространен и в центральных структурах, и на периферии медиатор ацетилхолин.
Химические медиаторы являются веществами, синтезируемыми в цитоплазме нейронов. Нейроны могут:
1) синтезировать один медиатор;
2) транспортировать медиатор по аксону с помощью микротрубочек;
3) упаковывать медиатор в гранулы;
4) хранить эти гранулы в терминалях аксонов;
5) освобождать медиатор при возбуждении;
6) захватывать назад в терминаль аксона медиатор или его фрагменты;
7) восстановить гранулы и вновь заполнить их медиатором.
Медиатор выделяется в синаптическую щель размером 10-50 мкм, и взаимодействует с мембранным рецептором постсинаптической мембраны.
В синаптической щели возможно разрушение медиатора специфическими для каждого медиатора ферментами, метаболиты возвращаются в нейрон для ресинтеза. Такими ферментами могут быть ацетилхолинэстераза – фермент для разрушения ацетилхолина, моноаминооксидаза (МАО) – для разрушения норадреналина или дофамина. Возможен обратный захват не разрушенного медиатора терминалью аксона.
Постсинаптическая мембранаимеетмембранные рецепторы– сложные комплексы интегральных белков, пронизывающих клеточную мембрану, как правило, состоящие из нескольких субъединиц. В мембранных рецепторах одна часть способна распознавать собственный медиатор и связываться с ним, а вторая – представлена или ионным каналом, или системой специальных мембранных ферментов, которые называются внутриклеточными посредниками.
Если взаимодействие медиатора с рецептором приводит к активации ионных каналов, а результатом является изменение потенциала мембраны, то такие рецепторы называются ионотропными. Если запускается система вторичных внутриклеточных посредников, происходит изменение активности ферментов и темпа метаболических процессов (гликолиз, липолиз), то такие рецепторы называются метаботропными. Вернитесь к рисунку 9, на котором представлена активация натриевого канала при взаимодействии ацетилхолина с холинорецептором – это пример ионотропного рецептора.

Для выделения медиаторанеобходимо сочетание двух событий: 1) волна деполяризации (ПД), достигшая терминали аксона, эта волна обусловлена активацией быстрых натриевых каналов и поступлением ионов натрия в клетку и 2) открытие медленных, потенциал-зависимыхкальциевыхканалов пресинаптической мембраны. Поступление кальция в клетку (по градиенту концентрации) стимулирует освобождение гранул с медиатором. Гранулы выделяются путем экзоцитоза, поэтому медиатор появляется в синаптической щели не отдельными молекулами, а квантами, примерно из нескольких тысяч молекул.
Рисунок 20. Выделение медиатора и взаимодействие с рецепторами
На рисунке 21 представлены этапы выделения медиатора:
Активация натриевых каналов, ток натрия в терминаль аксона (в клетку) по градиенту концентрации и деполяризация пресинаптической мембраны
Активация потенциалзависимых медленных кальциевых каналов и поступление ионов кальция в клетку по градиенту концентрации. Ионы кальция обеспечивают освобождение медиатора.
Диффузия медиатора к постсинаптической мембране и взаимодействие со специфическими мембранными рецепторами.
Судьба медиатора в синаптической щели
Возвращение метаболитов в терминаль аксона
Разрушение медиатора специфическими ферментами
Обратный захват медиатора

Рисунок 21. Этапы выделения и разрушения медиатора
Результат взаимодействия медиатора с мембранными рецепторами постсинаптической мембраны приводит к изменениям проницаемости этой мембраны для ионов путем активации или инактивации ионных каналов. На постсинаптической мембране могут открываться (или закрываться) натриевые, кальциевые, калиевые или хлорные каналы. Результатом этого процесса будет движение ионов по градиенту концентрации и изменение мембранного потенциала постсинаптической мембраны. В случае активации натриевых каналов ионы натрия по градиенту концентрации будут поступать в клетку и произойдет деполяризация постсинаптической мембраны. Если деполяризующий ток ионов велик (натриевые каналы), на постсинаптической мембране возникает импульс возбуждения – потенциал действия. Чаще же на постсинаптической мембране возникает лишь незначительная, не достигающая порогового уровня деполяризация, локальный ответ (ЛО). В зависимости от расположения синапса этот ЛО называется или ВПСП (возбуждающий постсинаптический потенциал в центральных синапсах) или ПКП (потенциал концевой пластинки в нервномышечных синапсах).

Рисунок 22. Изменение потенциалов пресинаптической и постсинаптической мембраны
1 – пресинаптическая мембрана, 2 – постсинаптическая мембрана
Синапсы могут быть как возбуждающими, так и тормозными, это зависит от того какой медиатор синтезируется нейроном, с какими рецепторами на постсинаптической мембране он взаимодействует и какие в результате этого взаимодействия открываются каналы. Если на постсинаптической мембране открываются натриевые каналы, то на постсинаптической мембране формируется ВПСП, а если открываются калиевые каналы, то ионы кали по градиенту концентрации будут выходить их клетки. В этом случае потенциал постсинаптической мембраны станет еще более отрицательным, мембрана окажется гиперполяризована. Поскольку при гиперполяризации возбудимость снижается, то такое изменение мембранного потенциала получило название тормозного постсинаптического потенциала (ТПСП). Сравните эти изменения потенциала постсинаптической мембраны на рисунке 23.
Возникновение потенциала действия в постсинаптическом нейроне. Постоянное взаимодействие возбуждающей и тормозящей активности на постсинаптическом нейроне приводит к изменениям МП, представляющим собой алгебраическую сумму деполяризующих и гиперполяризующих влияний. В этом отношении дендритная зона действует как определённого вида интегратор. Участком нервной клетки с самым низким порогом для генерации ПД является аксонный холмик, содержащий в своей мембране в 7 раз больше потенциалзависимых Na+ ‑ каналов, чем дендритная зона, и соответственно может генерировать ПД с большей лёгкостью в том случае, если суммарное изменение потенциала мембраны достигает критического уровня деполяризации. Возникший ПД распространяется в двух направлениях: вдоль аксона и по телу нейрона. В теле нейрона и дендритах ПД затухает, так как в этих отделах клетки мало потенциалзависимых Na+ ‑каналов. В терминальном расширении аксона ПД приводит к выделению медиатора в синаптическую щель.


Рисунок 23. Возбуждающие и тормозные синапсы
Синапсы можно классифицировать по локализации: на центральные и периферические. Центральные – это синапсы между нейронами, они отличаются местом взаимодействия терминали пресинаптического нейрона с постсинаптическим и могут быть аксо-соматическими, аксо-аксональными, аксо-дендритическими и дендро-дендритическими, кроме того, бывают аксо-вазальные синапсы, в которых медиатор выделяется в кровь. Периферические – это синапсы нервно-мышечные и нервно-секреторные, которые могут быть и соматическими и вегетативными: симпатическими и парасимпатическими. В периферических синапсах представлено два основных медиатора: ацетилхолин и норадреналин.
Очень широкие возможности модулировать (видоизменить) работу синаптического контакта открывает наличие в ЦНС аксо-аксональных синапсов. Такие синапсы, в которых выделяются другие медиаторы, могут или «помочь» выделению медиатора по основному пути (рис. 24Б), или «помешать», ослабить выделение медиатора (рис.24 А).


А

Рисунок 24. Взаимодействие синапсов
Мембранные рецепторы тоже можно классифицировать и по веществу, с которым они взаимодействуют, и по эффекту такого взаимодействия. Рецепторы, взаимодействующие с ацетилхолином (АХ) называются холинорецепторами. В функциональном отношении они разделяются на две группы: М- и Н-холинорецепторы. М — чувствительные к мускарину, Н — чувствительные к никотину. В синапсах скелетных мышц присутствуют только Н-холинорецепторы, а в гладких мышцах внутренних органов — преимущественно М-холинорецепторы. Рецепторы, взаимодействующие с норадреналином называются адренорецепторами, и делятся на альфа- и бета- адренорецепторы. В постсинаптической мембране гладкомышечных клеток внутренних органов и кровеносных сосудов часто соседствуют оба вида адренорецепторов. Действие норадреналина является деполяризующим, если он взаимодействует с альфа-адренорецепторами и гиперполяризующим при взаимодействии с бета-адренорецепторами (таблица 6). Холин- и адрено — реактивные структуры находятся во всех внутренних органах, железах внутренней и внешней секреции, скелетной и гладкой мускулатуре, вегетативных ганглиях и ЦНС.
Обратите внимание на то, что чувствительность адренорецепторов к адреналину и норадреналину различна, поэтому можно наблюдать несколько различные эффекты при активации симпатической нервной системы и при повышении в крови адреналина.