
Процесс возникновения потенциала действия в возбудимых тканях
В нервных волокнах сигналы передаются с помощью потенциалов действия, которые представляют собой быстрые изменения мембранного потенциала, быстро распространяющиеся вдоль мембраны нервного волокна. Каждый потенциал действия начинается со стремительного сдвига потенциала покоя от нормального отрицательного значения до положительной величины, затем он почти так же быстро возвращается к отрицательному потенциалу. При проведении нервного сигнала потенциал действия движется вдоль нервного волокна вплоть до его окончания.
На рисунке показаны изменения, возникающие на мембране во время потенциала действия, с переносом положительных зарядов внутрь волокна вначале и возвращением положительных зарядов наружу в конце. В нижней части рисунка графически представлены последовательные изменения мембранного потенциала в течение нескольких 1/10000 сек, иллюстрирующие взрывное начало потенциала действия и почти столь же быстрое восстановление.
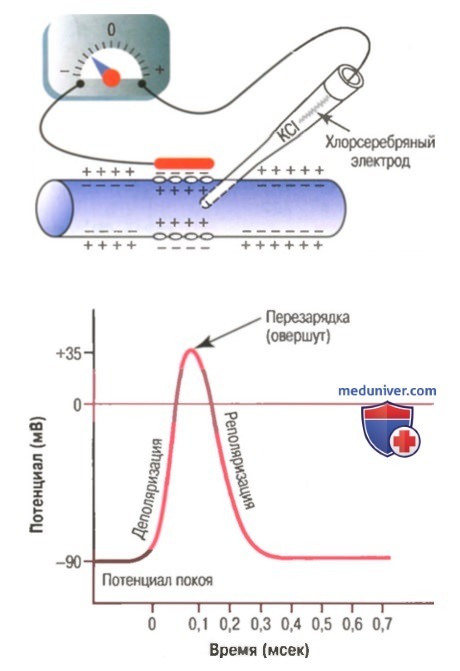
Типичный потенциал действия, зарегистрированный с помощью метода, представленного в верхней части рисунка.
Стадия покоя. Эта стадия представлена мембранным потенциалом покоя, который предшествует потенциалу действия. Мембрана во время этой стадии поляризована в связи с наличием отрицательного мембранного потенциала, равного -90 мВ.
Фаза деполяризации. В это время мембрана внезапно становится высокопроницаемой для ионов натрия, позволяя огромному числу положительно заряженных ионов натрия диффундировать внутрь аксона. Нормальное поляризованное состояние в -90 мВ немедленно нейтрализуется поступающими внутрь положительно заряженными ионами натрия, в результате потенциал стремительно нарастает в положительном направлении. Этот процесс называют деполяризацией, В крупных нервных волокнах значительный избыток входящих внутрь положительных ионов натрия обычно приводит к тому, что мембранный потенциал «проскакивает» за пределы нулевого уровня, становясь слегка положительным. В некоторых более мелких волокнах, как и в большинстве нейронов центральной нервной системы, потенциал достигает нулевого уровня, не «перескакивая» его.
Фаза реполяризации. В течение нескольких долей миллисекунды после резкого повышения проницаемости мембраны для ионов натрия, натриевые каналы начинают закрываться, а калиевые — открываться. В результате быстрая диффузия ионов калия наружу восстанавливает нормальный отрицательный мембранный потенциал покоя. Этот процесс называют реполя-ризацией мембраны.
Для более полного понимания факторов, являющихся причиной деполяризации и реполяризации, необходимо изучить особенности двух других типов транспортных каналов в мембране нервного волокна: электроуправляемых натриевых и калиевых каналов.
Электроупавляемые натриевые и калиевые каналы. Необходимым участником процессов деполяризации и реполяризации во время развития потенциала действия в мембране нервного волокна является электроуправляемый натриевый канал. Электроуправляемый калиевый канал также играет важную роль в увеличении скорости реполяризации мембраны. Оба типа электроуправляемых каналов существуют дополнительно к Na + /K + -насосу и каналам К + /Na + -утечки.
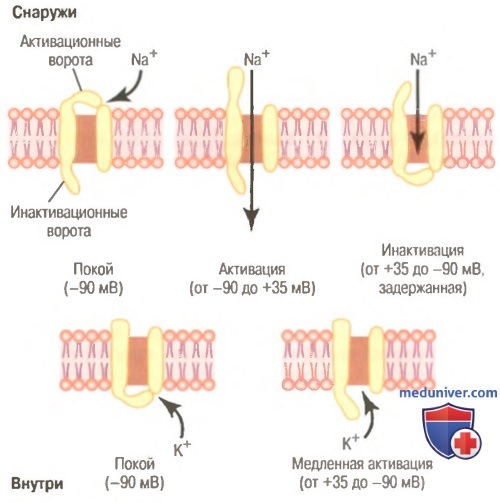
Типичные изменения состояния электроуправляемых натриевых (вверху) и калиевых (внизу) каналов при изменении мембранного потенциала от нормального отрицательного значения до положительных величин.
Показана последовательная активация и инактивация натриевых каналов и задержанная активация калиевых каналов.
Электроуправляемый натриевый канал. В верхней части рисунка показан электроуправляемый натриевый канал в трех различных состояниях. Этот канал имеет двое ворот: одни вблизи наружной части канала, которые называют активационными воротами, другие — у внутренней части канала, которые называют инактивационными воротами. В верхней левой части рисунка изображено состояние этих ворот в покое, когда мембранный потенциал покоя равен -90 мВ. В этих условиях активационные ворота закрыты и препятствуют поступлению ионов натрия внутрь волокна.
Активация натриевого канала. Когда мембранный потенциал покоя смещается в направлении менее отрицательных значений, поднимаясь от -90 мВ в сторону нуля, на определенном уровне (обычно между -70 и -50 мВ) происходит внезапное конформационное изменение актива-ционных ворот, в результате они переходят в полностью открытое состояние. Это состояние называют активированным состоянием канала, при котором ионы натрия могут свободно входить через него внутрь волокна; при этом натриевая проницаемость мембраны возрастает в диапазоне от 500 до 5000 раз.
Читайте также: Rust как добыть ткань
Инактивация натриевого канала. В верхней правой части рисунке показано третье состояние натриевого канала. Увеличение потенциала, открывающее активационные ворота, закрывает инактивационные ворота. Однако инактивационные ворота закрываются в течение нескольких десятых долей миллисекунды после открытия активационных ворот. Это значит, что конформационное изменение, приводящее к закрытию инактивационных ворот, — процесс более медленный, чем конформационное изменение, открывающее активационные ворота. В результате через несколько десятых долей миллисекунды после открытия натриевого канала инактивационные ворота закрываются, и ионы натрия не могут более проникать внутрь волокна. С этого момента мембранный потенциал начинает возвращаться к уровню покоя, т.е. начинается процесс реполяризации.
Существует другая важная характеристикая процесса инактивации натриевого канала: инактивационные ворота не открываются повторно до тех пор, пока мембранный потенциал не вернется к значению, равному или близкому к уровню исходного потенциала покоя. В связи с этим повторное открытие натриевых каналов обычно невозможно без предварительной реполяризации нервного волокна.
Видео физиология возбуждения тканей (потенциал покоя, потенциал действия) — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в оглавление раздела «Физиология человека.»
Процесс возникновения потенциала действия в возбудимых тканях
• Потенциал действия обеспечивает клеткам способность быстро взаимодействовать друг с другом
• Ключевыми элементами потенциала действия являются потоки ионов Na+, К+ и Са2+
• Деполяризация мембраны обеспечивается потоком в клетки ионов Na+ через потенциал-зависимые Na+-каналы
• Реполяризация возникает за счет транспорта ионов К+ через несколько различных К+-каналов
• Электрическая активность органов выражается как сумма векторов потенциала действия
• Нарушения генерации или распространения потенциала действия могут предрасполагать к развитию аритмий или эпилепсии
Нейроны, мышечные и эндокринные клетки относятся к числу возбудимых клеток, поскольку они могут создавать электрические сигналы и/или отвечать на них. В этих клетках происходят быстрые обратимые изменения мембранного потенциала, которые переводятся в электрические сигналы, например в нервный импульс, распространяющийся по аксону нейрона, или в сигнал, который приводит к мышечному сокращению. Такой электрический сигнал называется потенциал действия.
Процесс восприятия в головном мозгу включает обработку огромного количества потенциалов действия. Эти потенциалы действия могут возникать до появления стимула, одновременно с ним, или после него, и имеют различные пространственные и временные характеристики. В клетках скелетных мышц и миокарда потенциал действия играет критическую роль в инициации и координации процесса их синхронного сокращения. Амплитуда и продолжительность потенциала действия измеряются электрофизиологическими методами. В нейронах продолжительность потенциала действия составляет несколько миллисекунд, а в клетках миокарда — сотни миллисекунд.
Более высокая продолжительность потенциала действия в клетках миокарда необходима для координации процессов активации в миллионах мышечных клеток, ответственных за сердечные сокращения. Распространение потенциала действия по мембране происходит со скоростью несколько метров в секунду. Этим обеспечивается быстрая взаимосвязь на значительном расстоянии между клетками. Такая связь является основой для реализации сложных физиологических функций головного мозга и сердца.
Читайте также: Ткань антивандальная названия цветов
Ключевую роль в генерации потенциала действия играет трансмембранный потенциал. В состоянии покоя в клетке поддерживается отрицательное значение мембранного потенциала, т. е. внутренняя ее среда по отношению к внешнему окружению заряжена несколько более отрицательно.
Мембранный потенциал покоя поддерживается, главным образом, за счет действия Na+/K+. АТФаз, которые нагнетают в клетку три иона Na+ на каждые два иона К+, транспортируемые из клетки, а также за счет функционирования К+-каналов.
Около пятидесяти лет назад на основании электрофизиологических данных была предложена модель механизма возникновения потенциала действия. Модель предусматривает два ключевых момента, определяющих возникновение потенциала действия:
• наличие обратимых скоординированных изменений в селективной проводимости клеточной мембраны для ионов Na+ или К+;
• эти изменения проводимости зависят от мембранного потенциала.
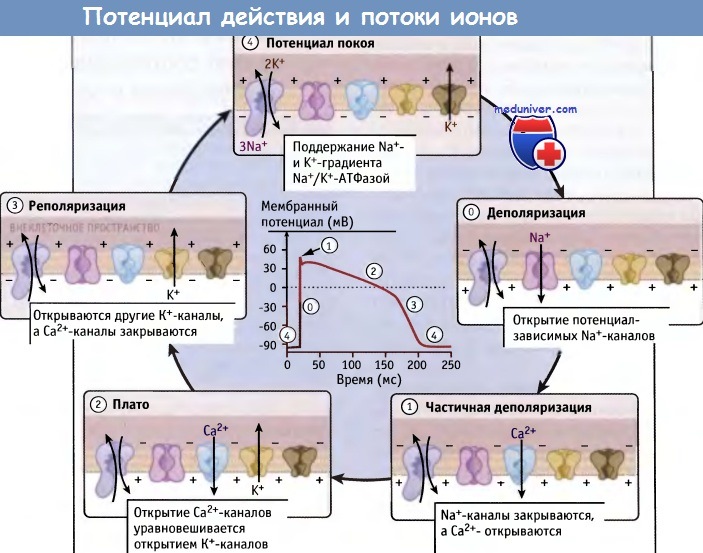
Генерация и распространение потенциала действия обусловлены потоками ионов.
Потенциал действия клеток миокарда подразделяется на пять фаз.
Каждая фаза связана с открытием и/или с закрытием определенных ионных каналов.
Разные типы К+-каналов, которые открываются и закрываются в различные фазы потенциала действия, неспецифичны для данной фазы и показаны одинаковым образом.
Во время генерации и распространения потенциала действия функционирует Na+/K+-ATФазa,
однако скорость ее работы меняется по мере изменения мембранного потенциала.
Потенциал действия возникает за счет скоординированных процессов активации и инактивации ионных каналов нескольких различных типов. Возникновение электрических сигналов в мембране становится возможным при реализации сенсорной функции потенциал-зависимых ионных каналов и их реакции на изменения мембранного потенциала. Как показано для клеток миокарда, открытие и закрытие различных типов ионных каналов происходит последовательным образом в различных фазах потенциала действия.
Быстрые изменения мембранного потенциала обусловлены локальными трансмембранными изменениями концентрации ионов, которые почти не влияют на общую их концентрацию в клетке.
Потенциал действия возникает при быстром скачке заряда (фаза 0), который инициируется открытием потенциал-зависимых Na+-каналов и обеспечивает быстрый поток в клетку ионов натрия в направлении их концентрационного градиента. В результате транспорта ионов натрия происходит деполяризация мембраны, и внутриклеточная среда приобретает более положительный заряд по сравнению с внеклеточным окружением.
Через несколько миллисекунд, по мере того как происходит быстрая инактивация Na+-каналов, деполяризация прекращается, и наступает ранняя реполяризация (фаза 1). Когда в клетках миокарда происходит закрытие потенциал-зависимых Na+-каналов, открываются потенциал-зависимые Са2+-каналы и выводящие К+-каналы. Эти каналы активируются по мере роста положительного мембранного потенциала, который происходит при деполяризации мембраны. Устанавливается новая величина мембранного потенциала. Эта величина определяется тонким балансом между токами деполяризации и реполяризации мембраны в фазе 2.
Для того чтобы Na+-каналы снова приобрели способность к активации, они должны выйти из фазы инактивации, а для этого необходима реполяризация мембранного потенциала. Небольшая часть Na+-каналов не инактивируется и обеспечивает небольшой, но постоянный поток ионов, который наряду с поддерживающим транспортом Са2+ в клетку способствует продолжительности пребывания клеток миокарда в деполяризованном состоянии. По сравнению с потенциалом действия нейронов, потенциал действия клеток миокарда проявляется в течение более длительного промежутка времени. Длительный потенциал действия необходим для того, чтобы обеспечить достаточно времени для высвобождения внутриклеточного кальция, необходимого для мышечного сокращения и предотвращения при этом аберрантной мембранной деполяризации.
В клетках крупных животных и человека фазы 1 и 2 потенциала действия разделены небольшим пиком. В фазе 2 (плато-фазе) при последовательной активации нескольких различных К+-каналов из клетки выходят ионы К+. При этом их убыль быстро уравновешивается положительно заряженными ионами, поступающими в клетку по Na+- и Са2+-каналам. Более того, Na+/Ca2+ обменник вытесняет ионы Са2+ из цитозоля, создавая деполяризующий поток, направленный внутрь клетки. Вместе с тем, Na+/К+-АТФаза продолжает вытеснять из клетки три иона Na+ в обмен на два иона К+, что сдвигает мембранный потенциал в сторону реполяризации (фаза 3).
Читайте также: Выкройка балалайки из ткани
Попеременное открытие и закрытие различных ионных каналов снижают потенциал действия, и в клетке устанавливается отрицательный мембранный потенциал покоя (фаза 4). Для прекращения эффекта потенциала действия необходима высокая скорость транспорта ионов. Это достигается с помощью К+-каналов, которые к тому же проявляют высокую селективность. Например, для того чтобы быстро снять эффект потенциала действия в нейронах, из клетки за миллисекунду выходят миллионы ионов К+
В различных типах клеток генерация и передача электрического сигнала обеспечивается участием большого количества потенциал-зависимых К+-каналов. Например, входные выпрямляющие К+-каналы необходимы для установления стабильного мембранного потенциала покоя и продолжительной плато-фазы в клетках миокарда. При положительном мембранном потенциале эти каналы в основном закрыты, что обеспечивает большую продолжительность процесса мембранной деполяризации. Изначально, в клетках миокарда, когда значение мембранного потенциала меняется от -40 мВ в положительную сторону, отмечается лишь минимальный поток ионов К+. Это поддерживает влияние деполяризующих потоков натрия и кальция на продолжительность потенциала действия до момента активации К+-канала входящего выпрямления, в результате функционирования которого мембранный потенциал возвращается к состоянию покоя.
Суммарная электрическая активность, которая складывается из индивидуальных потенциалов действия всех нейронов мозга, всех клеток определенной группы мышц, или всех клеток миокарда, после амплификации может быть зарегистрирована в виде электроэнцефалограммы (ЭЭГ), электромиограммы (ЭМГ) или электрокардиограммы (ЭКГ) соответственно. Процедура регистрации изменений электрической активности используется для контроля над развитием различных патологических состояний: эпилепсии, миотоний или аритмий. Развитие этих патологических состояний может явиться следствием мутаций, влияющих на функционирование определенных типов ионных каналов.
Мутации, возникающие в генах, кодирующих белки потенциал-зависимых Na+-, К+- или Са2+-каналов, обусловливают нарушения функций головного мозга и сердца. Например, мутации в гене SCN5A, кодирующем белок потенциал-зависимого натриевого канала клеток миокарда, вызывают некоторые болезни сердца. В результате некоторых типов мутаций в этом гене развивается синдром удлиненного QT, при котором неполная инактивация Na+-каналов увеличивает продолжительность потенциала действия. Наряду с этим, мутации в гене одного из белков Cavl,2 канала, который определяет длительность плато-фазы потенциала действия, приводят к развитию синдрома удлиненного QT и аритмий.
Последние вызывают задержку реполяризации сердца и увеличивают у больных риск внезапной смерти. Другие мутации, затрагивающие белки потенциал-зависимых Na+-каналов, приводят к развитию различных болезней сердца, параличам скелетных мышц или к наследственным формам эпилепсии. Примером мутации в белках потенциал-зависимого К+-канала, связанной с болезнью сердца, является мутация в гене HERG. Она увеличивает скорость инактивации канала, что снижает поток ионов К+ из клетки и замедляет фазу реполяризации потенциала действия. Таким образом, эта мутация увеличивает продолжительность потенциала действия, подобно тому как это происходит при мутациях в генах, кодирующих потенциал-зависимые Na+-каналы.
Видео физиология возбуждения тканей (потенциал покоя, потенциал действия) — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021