
ЭЛЕКТРОПРОВОДНОСТЬ БИОЛОГИЧЕСКИХ СИСТЕМ — количественная характеристика способности живых объектов (тканей) проводить электрический ток. Электропроводность обратно пропорциональна величине электрического сопротивления системы.
Измерение электропроводимости биологических систем используют для получения информации о функциональном состоянии биол, тканей, для выявления воспалительных процессов, изменения проницаемости клеточных мембран и стенок сосудов при патологии или действии на организм физических, химических и других факторов (см. Проницаемость). Измерение электропроводимости биологических систем лежит в основе многих методов оценки кровенаполнения сосудов органов и тканей (см. Реография).
При подаче разности потенциалов (U) через объект течет электрический ток силой (I), величина которой пропорциональна электропроводности (G): I=GU. Величина электропроводности зависит от количества свободных электрических зарядов и их подвижности. Чем больше количество зарядов и их подвижность, тем больше электропроводность. В клетке основными свободными зарядами являются ионы калия и органические анноны, а снаружи клетки, в межклеточных пространствах и тканевых жидкостях — ионы натрия и хлора. В биологических жидкостях (кровь, спинномозговая жидкость, моча и т. д.) электропроводность пропорциональна содержанию в них свободных ионов. Связанные заряды (ионогенные группы белков, липидов, углеводов), перемещение которых ограничено, и крупные ионы с малой подвижностью не оказывают существенного влияния на величину электропроводимости биологических систем.
Электропроводность или сопротивление клеток, тканей, органов и целых организмов измеряют при пропускании через них постоянного или переменного синусоидальной формы тока, частота которого может изменяться от долей герца до 10 10 гц. При измерениях на переменном токе с круговой частотой ω (ω = 2πf, где f — частота в гц) общее сопротивление системы, или импеданс (см.), зависит от наличия границ раздела в системе, на которых может происходить накопление зарядов — поляризация (см.). Свойства границ раздела (в биологическом объекте это главным образом различные мембраны) могут быть описаны, если ввести понятие емкости С, сопротивление которой Хс (реактивное сопротивление в отличие от R — активного сопротивления) зависит от частоты, на которой производится измерение: Xс = 1/(ωC).
Общее сопротивление (Z) равно сумме сопротивлений реактивного Хc и активного R, если R и С соединены последовательно; при параллельном соединении R и С общее сопротивление рассчитывается по формуле:

Для измерения электрических характеристик биол. объекта применяют металлические или угольные электроды (см.), которые прикладывают к объекту с помощью жидкостного контакта — тонкого слоя хорошо проводящей жидкости, чаще всего — физиологического раствора. В ряде случаев, напр. при измерении электропроводности цитоплазматических мембран, один из электродов вводится внутрь клетки, а другой подводится к клетке снаружи (см. Микроэлектродный метод исследования). При измерении электропроводимости биологических систем на постоянном и переменном токах низкой частоты важно учитывать величину поляризации электродов, поскольку за счет электродной поляризации истинные электрические параметры биол, объекта могут значительно отличаться от измеренных. Величина поляризации электродов определяется плотностью тока, которая зависит от приложенной к системе разности потенциалов, сопротивления системы, площади измерительных электродов. Чем больше площадь электродов, тем меньше плотность тока и тем меньше искажения, вносимые в измерения электродами. Поэтому для уменьшения плотности тока используют электроды с большой эффективной поверхностью, в частности платиновые электроды, покрытые платиновой чернью (их губчатое покрытие увеличивает эффективную поверхность электродов в 100 — 1000 раз). Возможно применение и так называемых неполяризующихся электродов (например, каломельных, хлорсеребряных).
Для облегчения интерпретации получаемых результатов биологическую систему (ткани, суспензии клеток) часто представляют в виде модели — электрической схемы, состоящей из набора активных сопротивлений и емкостей, являющихся как бы эквивалентами биологических структур клеток или тканей, участвующих в проведении электрического тока.
Измерение электропроводимости биологических систем на постоянном токе из-за высокой степени поляризации мембран и электродов крайне затруднено. На низких частотах переменного тока большая часть тока протекает по межклеточным промежуткам. При увеличении частоты электрического тока реактивное сопротивление емкости падает, поляризационные явления уменьшаются. Зависимость сопротивления и емкости объекта от частоты получила название дисперсии (см.). На высоких частотах общее сопротивление системы зависит только от активных сопротивлений межклеточных пространств и цитоплазмы.
В медицине и биологии электропроводимость биологических систем чаще всего исследуют в области так называемой β-дисперсии, которая наблюдается в диапазоне частот 10 2 —10 8 гц и определяется поляризацией границ раздела и неоднородностью структуры объекта. Измерения электропроводимости биологических систем показали, что по мере повышения частоты электропроводность возрастает, достигая предельной величины. При переживании и отмирании ткани возрастает электропроводимость биологических систем на низких частотах. Это связано с тем, что при отмирании ткани растет проницаемость мембран для ионов, и они уже не являются границей, на которой может происходить поляризация. Основываясь на способности живой ткани к поляризации, Б. Н. Тарусов предложил в качестве критерия оценки жизнеспособности ткани использовать коэффициент К, численно равный отношению Rн/Rв где Rн и Rв — сопротивления ткани, измеренные соответственно на низкой и высокой частотах; при отмирании ткани он стремится к единице. Выбор частот для расчета К определяется диапазоном β-дисперсии: низкой частоте соответствует частота начала β-дисперсии, высокой — частота, при которой электропроводимость биологических систем достигает максимальной величины. Например, для мышечной ткани — это 10 3 и 10 6 гц, клеток крови и жировой ткани — 10 4 и 10 7 гц, кожи — 10 2 и 10 4 гц и т. д. На высоких частотах, когда активное и общее сопротивления не зависят от частоты, возможно исследование относительных изменений числа ионов в биол. системе, связанных с нарушением водно-солевого обмена.
Читайте также: Обивка стула расчет ткани
Сопоставляя Данные, полученные при измерении на низких и высоких частотах, можно вычислить объем и ионную проводимость межклеточных пространств и цитоплазмы клеток, проницаемость мембран для ионов, емкостные характеристики мембраны. Если измерения проводятся в системе, где межклеточные пространства занимают достаточно большой объем (более 20—30%), например при измерениях электропроводности крови, то для вычисления параметров дисперсной фазы (эритроцитов) используют специальные формулы. Частоты, на которых наблюдается дисперсия, зависят от величины клеток и объема межклеточных пространств. Так, дисперсия электропроводимости биологических систем для клеток крови начинается на частотах порядка нескольких десятков килогерц, для мышечной ткани — несколько килогерц, жировой — сотен килогерц. При исследовании электрических характеристик плазматических мембран клеток дисперсия обнаруживается на частотах порядка нескольких десятков герц. Электрические характеристики тканей и органов на низких частотах зависят от неоднородности расположения клеток и межклеточных пространств и соотношения их объемов. Этот факт используется в реографии и реоэнцефалографии (см.) при исследовании изменений кровенаполнения ткани и эластических свойств стенок сосудов. Измерение электропроводимости биологических систем на низких частотах позволяет оценить изменения объема межклеточных пространств, в частности при развитии воспаления (см.). Так, на первых стадиях воспалительного процесса структура клеток изменяется незначительно, и импеданс клеток сохраняет свою величину. По мере набухания клеток и уменьшения объема межклеточных пространств происходит увеличение общего сопротивления системы. На более поздних стадиях развития воспаления импеданс системы уменьшается за счет возрастания проницаемости мембран для различных ионов.
Таким образом, измерение электропроводимости биологических систем или импеданса, особенно в широком диапазоне частот, может быть использовано при исследовании проницаемости клеточных мембран и других границ раздела в клетках, тканях, органах, а стандартизация (измерение удельных величин) дает возможность сравнивать данные, полученные разными исследователями. Возбуждение, изменение интенсивности метаболизма и других функций клеток приводят к изменению электропроводимости биологических систем. Методы измерения электропроводимости биологических систем используют для исследования влияния на биологические объекты различных факторов: работы (увеличение интенсивности метаболизма приводит к увеличению проницаемости мембран); психогенных (изменяется проницаемость кожи за счет работы потовых желез); физических (радиация, ультрафиолетовое излучение, температура и др.) и химических (кислоты, щелочи, спирты и др.), обычно сопровождаемых ростом проницаемости. Изменение проницаемости мембран часто зависит от дозы или концентрации действующего вещества. Так, соли меди в малых концентрациях уменьшают проницаемость мембран мышечных клеток кожи лягушки, а в концентрациях более 10 -3 М — увеличивают. Исследование электрических свойств возбудимых тканей способствовало изучению механизма проведения возбуждения по нерву п мышце. На основании измерений активного сопротивления, емкости и их дисперсии была вычислена статическая емкость клеточной мембраны (около 1 мкф/см 2 ) и впервые определена толщина ее липидного бислоя. Было найдено, что удельное сопротивление аксоплазмы и миоплазмы всего в 2—3 раза выше сопротивления внеклеточной жидкости, тогда как сопротивление мембраны выше в десятки тысяч раз. Эти данные послужили основанием для возникновения представления о «кабельной» структуре волокна. Установлены временные соотношения между изменением проницаемости мембраны для ионов и развитием потенциала действия — «импедансный спайк» (см. Биоэлектрические потенциалы, Нервный импульс). Исследование электропроводимости биологических систем может быть использовано для оценки состояния тканей при их консервации, а также эффективности действия биологически активных веществ на модельные системы. В ряде случаев проницаемость биол. мембран для ионов сопряжена с их проницаемостью для незаряженных частиц— сахаров, аминокислот и других соединений. Поэтому измерение электропроводимости биологических систем может оказаться полезным при изучении проницаемости мембран и для неэлектролитов. Исследование электпроводимости биологических систем может найти применение и в биотехнологии для оценки оптимальности среды и условий культивирования клеток.
Читайте также: Как правильно кроить ворсовые ткани
Библиогр.: Андреев В. С. Кондуктометрические методы и приборы в биологии и медицине, М., 1973; Биофизика, под ред. Б. Н. Тарусова и О. Р. Колье, с. 186, М., 1968; Гречин В. Б. и Боровикова В. Н. Медленные неэлектрические процессы в оценке функционального состояния мозга человека, с. 22, Л, 1982; Гуревич М. И. и др. Импедансная реоплетизмография, Киев, 1982; Егоров Ю. В. и Кузнецова Г. Д. Мозг как объемный проводник, М., 1976; Слынько П. П. Основы низкочастотной кондуктометрии в биологии, М., 1972; Хассет Дж. Введение в психофизиологию, пер. с англ., с. 53, М., 1981; Электроника и кибернетика в биологии и медицине, пер. с англ., под ред. П. К. Анохина, с. 71, М., 1963; Schwan Н. P. Electrical properties of tissue and cell suspensions, Advanc, biol. med. Phys., v. 5, p. 147, 1957.
Башкирский государственный медицинский университет
Нормальная физиология
Теория
Биоэлектрические явления в живых тканях. Потенциал покоя
В 1949-1952 гг. Ходжкин, Хаксли и Катц модифицировали существовавшие ранее теории и экспериментально обосновали основные положения мембранно-ионной теории .
Согласно этой теории возникновение МП (мембранного потенциала) обусловлено:
- Разной концентрацией ионов калия, натрия, кальция и хлора внутри клетки и во внеклеточной среде.
- Разной проницаемостью мембраны клеток для этих ионов в покое и при возбуждении.
Состояние покоя — это такое состояние, когда отсутствует раздражающее действие на организм. Это состояние относительное, т.к. абсолютный покой не совместим с жизнью.
Деятельное состояние — это изменение состояния организма, возникающее под действием факторов внешней среды.
Раздражимость — это способность организма отвечать на действие раздражающих факторов изменением его структурных и функциональных свойств.
Возбудимость
Возбудимость — способность ткани отвечать на раздражение специализированной реакцией — возбуждением. Возбудимость является одной из форм раздражимости.
Возбудимыми тканями называются такие ткани, клетки которых способны в ответ на раздражение генерировать специфическую реакцию — возбуждение .
Возбуждение — это специализированная ответная реакция живого объекта на действие раздражителя, проявляющаяся в изменениях его определенных параметров.
К возбудимым тканям относятся:
Общие признаки возбуждения (присущи всем возбудимым тканям):
- Изменение уровня обменных процессов в тканях;
- Выделение различных видов энергии — тепловой, электрической.
Специфические признаки возбуждения (характерны для определенного вида тканей):
- Мышечная ткань — сокращение,
- Железистая — выделение секрета,
- Нервная — генерация и проведение нервного импульса.
- эпителиальная,
- соединительная ткани.
Физиологические свойства возбудимых тканей
Основными свойствами всех возбудимых тканей являются:
Лабильность (функциональная подвижность) — это способность ткани реагировать на различные воздействия с определенной скоростью, то есть способность воспроизводить определенную частоту раздражения.
Мерой лабильности является наибольшее число ответных реакций, которыми возбудимая ткань способна реагировать за единицу времени в соответствии с частотой приложенных к ней раздражений.
Проводимость — это способность ткани и клетки передавать возбуждение.
Раздражение — это процесс воздействия на живую ткань агентов внешней по отношению к этой ткани среды.
Раздражитель — это причина, способная вызвать ответную реакцию.
Классификация раздражителей
- физические,
- химические,
- физико-химические,
- биологические.
По биологическому значению раздражители делятся на 2 группы :
- Адекватные — раздражители, к воздействию которых ткани в процессе эволюции приспособлены в наибольшей степени.
- Неадекватные — раздражители, к воздействию которых возбудимые ткани специально не приспособлены.
Состав плазматической мембраны
- Липиды (в основном фосфолипиды),
- Белки (гликопротеины),
- Углеводы (мукополисахариды).
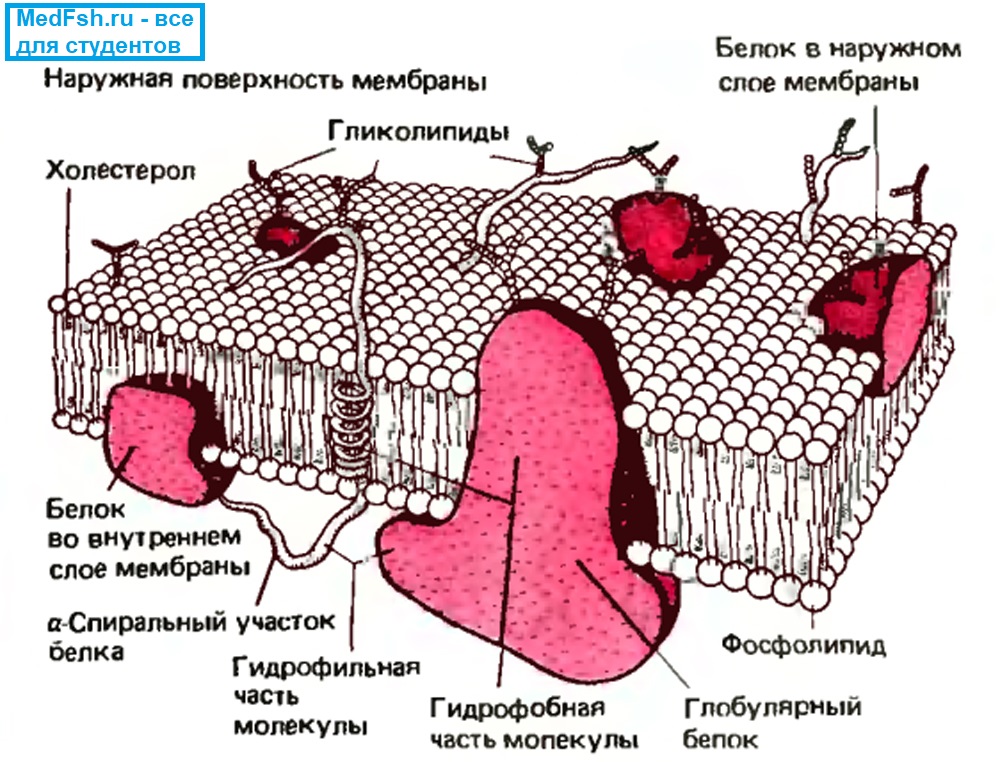
Липиды очень плотно упакованы в мембране, между ними нет практически никаких расстояний, поэтому мембрана плохо пропускает воду, практически непроницаема для ионов и других крупных молекул.
Белковые молекулы могут быть погружены в слой липидов с внеклеточной или с цитоплазматической стороны, либо могут целиком пронизывать мембрану.
Если белки прикреплены к поверхности мембраны, их называют периферическими. С внутренней стороны это будут белки-ферменты, а с наружной — белки-рецепторы.
Если белки пронизывают всю толщу мембраны клеток, то их называют интегральными или трансмембранными.
Такие белки образуют структуры, обеспечивающие движение ионов через мембрану.
Если белки образуют стенки поры, сквозь которую путем простой диффузии проходят ионы, то это ионные каналы.
Если трансмембранные белки перекачивают ионы против концентрационного и электрического градиентов, то это ионные насосы.
Все каналы, имеющиеся в живых тканях, можно разделить на 2 типа:
- первый тип — каналы покоя, которые спонтанно открываются и закрываются без всяких внешних воздействий;
- второй тип — gate-каналы (воротные каналы) — в покое они закрыты и открываются под действием раздражителей.
- неспецифические (каналы утечки, всегда открыты),
- специфические (селективные), обладающие способностью пропускать только определенные ионы при изменении заряда на мембране или действии химических веществ.
Читайте также: Тонкая шерстяная ткань это
Транспорт веществ
Транспорт частиц через каналы является жизненно важным для клеток процессом.
Обычно транспорт веществ делят на пассивный (без затраты энергии), т.е. транспорт веществ по концентрационному, осмотическому и электрохимическому градиентам и активный (с затратой энергии).
Различают первично- и вторично-активный транспорт
Первично-активный транспорт ионов обеспечивается специальными ионными насосами, осуществляется с затратой энергии АТФ, против концентрационного градиента, т.е. перенос веществ происходит из меньшей концентрации через мембрану в большую концентрацию.
- Это вид транспорта для переноса веществ (глюкозы, аминокислот и др.) через мембрану также против градиента, но без затраты энергии .
- Эти вещества проходят через мембрану с помощью специальных переносчиков (напр. ионов Na), на транспорт которых и затрачивается энергия, а эти вещества движутся как бы попутно.
Мембранный потенциал или потенциал покоя
В состоянии покоя между наружной и внутренней поверхностью мембраны возникает разность зарядов или потенциалов, которая в дальнейшем была названа мембранным потенциалом (МП) или потенциалом покоя (ПП) .
Положительные заряды концентрируются на наружной поверхности мембраны, а отрицательные заряды — на внутренней поверхности.
Мембранный потенциал измеряется в отрицательных значениях, т.к. внутренняя поверхность мембраны заряжена отрицательно. Его величина колеблется от -60 до -90 мВ у разных клеток.
Методы измерения мембранного потенциала
В зависимости от места приложения электродов :
- внеклеточный с помощью макроэлектродов,
- внутриклеточный с помощью микроэлектродов.
1. Исследование ПП с помощью макроэлектродов осуществляют, прикладывая один из них к неповрежденному , а другой — к поврежденному участку ткани.
Микроэлектрод представляет собой микропипетку диаметром 0,5 — 1 мкм, наполненную концентрированным солевым раствором (KCl). Внутри него может также находится неполяризующийся электрод — из серебра, золота или платины.
Второй электрод помещается во внеклеточную жидкость.
Оба электрода присоединяются к усилителю и осциллографу для регистрации потенциала. В момент прокалывания мембраны, осциллограф регистрирует появление отрицательного потенциала , соответствующего ПП.
1-ая причина — ионная асимметрия:
- Ионов K находится в клетке в 30-50 раз больше, чем снаружи;
- Ионов Na больше находится вне клетки, чем внутри нее в 8-10 раз;
- Ионов Ca во много раз больше вне клетки;
- Ионов Cl также больше во внеклеточной жидкости, чем внутри клетки в 50 раз;
- Внутри клетки находится больше органических анионов по сравнению с наружной поверхностью.
Таким образом, для этих ионов направление концентрационного градиента различно!
- Для K из клетки (из большей концентрации к меньшей);
- Для Na, Ca и Cl в клетку.
2-ая причина, обуславливающая поляризацию мембраны:
- различная проницаемость мембраны для различных ионов. В состоянии покоя мембрана в 25 раз более проницаема для ионов K, чем для Na, т.к. количество калиевых каналов на единицу площади мембраны намного больше, чем натриевых.
- Поскольку концентрация ионов K в цитоплазме намного выше, чем снаружи клетки, они начинают двигаться через канал и выходят из клетки.
- Ионы K несут положительные заряды, поэтому мембрана снаружи заряжается положительно.
Отрицательный заряд на внутренней поверхности мембраны обусловлен наличием органических анионов — крупномолекулярных соединений, которые заряжены отрицательно, и для которых мембрана непроницаема (глютамат, аспартат, органические фосфаты, сульфаты и др.)
Таким образов, на мембране формируется калиево-равновесный потенциал, т.к. уравновешиваются силы диффузии (выход K из клетки по градиенту концентрации) и электростатического взаимодействия (отталкивание выходящих ионов K положительным зарядом на наружной поверхности мембраны).
Калий является основным ионом, обеспечивающим формирование МП (ПП), что подтверждается формулой Нернста . По ней можно, зная концентрацию онов калия внутри и снаружи клетки, подсчитать величину ПП.

Натрий-калиевый насос или помпа
Для непрерывного активного переноса ионов Na и ионов K через мембрану клетки против концентрационного градиента существует Na-Л обменный насос. При каждом цикле он переносит 3 иона Na наружу и 2 иона K внутрь клетки.
Таким образом, МП представляет собой разность потенциалов между наружной и внутренней поверхностью мембраны клетки. Он является результатом разделения зарядов относительно клеточной мембраны , которое возникает за счет движения заряженных ионов (натрия и, преимущественно, калия) по концентрационным градиентам через ионные каналы, открытые в покое и за счет работы Na-K насоса .
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом